Approfondimenti |
|

 Il sistema nervoso è la sede dell'assunzione, dell'elaborazione e trasmissione delle informazioni; in altre parole è un sistema di regolazione delle funzioni coporee e "fa concorrenza" al sistema endocrino e immunitario. Il sistema nervoso è un complesso sistema elettrochimico, comnposto da oltre 1000 miliardi di cellule, che viaggia a 400 km all’ora e che assorbe il 20% dell’ossigeno del nostro organismo.
Il sistema nervoso è la sede dell'assunzione, dell'elaborazione e trasmissione delle informazioni; in altre parole è un sistema di regolazione delle funzioni coporee e "fa concorrenza" al sistema endocrino e immunitario. Il sistema nervoso è un complesso sistema elettrochimico, comnposto da oltre 1000 miliardi di cellule, che viaggia a 400 km all’ora e che assorbe il 20% dell’ossigeno del nostro organismo.
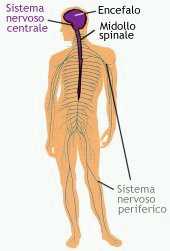
Encefalo e midollo spinale formano il sistema nervoso centrale o asse cerebrospinale (o nevrasse, neuroasse o tubo neurale) che è in grado di raccogliere, trasmettere e integrare le informazioni. Custodito all'interno della scatola scatola cranica (encefalo) e del canale vertebrale (midollo spinale), il sistema nervoso centrale, viene protetto ulteriormente e nutrito dal sistema delle meningi, dal liquor cerebrospinale e dalla propria vascolarizzazione (barriera emato-encegalica, emato-liquorale ed emato-retinica).
Il sistema nervoso periferico, è formato dai nervi cranici o cefalici e dai nervi spinali (formanti assieme i nervi encefalospinali) e dai relativi gangli. Il sistema nervoso periferico viene suddiviso in volontario o somatico e involontario o vegetativo (o viscerale) o autonomo.
 L'encefalo, con un peso di 1,3-1,5 kg, è il secondo organo più pesante del corpo dopo il fegato. A riposo, il 25% dell'energia metabolica viene utilizzata per rifornire il cervello (8-10% in più rispetto agli altri primati non umani).
Le strutture più antiche ("inferiori") si trovano in profondità (inferiormente e immediatamente sopra il forame occipitale), mentre le componenti più recenti ("superiori") si collocano sopra di esse.
L'encefalo, con un peso di 1,3-1,5 kg, è il secondo organo più pesante del corpo dopo il fegato. A riposo, il 25% dell'energia metabolica viene utilizzata per rifornire il cervello (8-10% in più rispetto agli altri primati non umani).
Le strutture più antiche ("inferiori") si trovano in profondità (inferiormente e immediatamente sopra il forame occipitale), mentre le componenti più recenti ("superiori") si collocano sopra di esse.
Il midollo allungato (bulbo o mielencefalo) è la parte più antica e la sua struttura si continua col midollo spinale (di cui ricorda la struttura metamerica). Nel midollo allungato i fasci motori, provenienti dalla corteccia cerebrale e diretti al midollo spinale, formano le piramidi, dove avviene la loro decussazione (incrocio a X). Il bulbo o midollo allungato, attraverso confini ben delineati, continua col ponte (ponte di Varolio) a cui si unisce il mesencefalo (o mesoencefalo), che assume una posizione mediana fra le regioni cerebrali antiche e recenti ("cervello intermedio"). Il mesencefalo è percorso da un sottile canale, l’acquedotto di Silvio (residuo della cavità mesencefalica embrionale). la sostanza grigia periacqueduttale media gli effetti analgesici (tramite la liberazione di endorfine). La porzione ventrale o corpo del mesoencefalo presenta i due peduncoli cerebrali (dx e sx), consistenti in colonne cilindriche di tessuto nervoso che mettono in comunicazione gli emisferi cerebrali con le altre strutture del sistema nervoso centrale. La porzione anteriore di ciascun peduncolo è caratterizzato da un nucleo intensamente pigmentato, detto sostanza nera del Sommering (substantia nigra) mentre posteriormente è presente il nucleo rosso. La sostanza nera di Sommering e il nucleo rosso sono importanti centri del sistema motorio extrapiramidale (la degenerazione e la conseguente decolorazione della substantia nigra di Sommering sono associate morbo di Parkinson). Sopra al nucleo rosso si trovano i nuclei dei due nervi oculomotori.
Dorsalmente il mesencefalo è caratterizzato da quattro collicoli (sporgenze rotondeggianti), i tubercoli quadrigemini, che creano una formazione caratteristica, definita lamina quadrigemina. I tubercoli corrispondono a nuclei di sostanza grigia appartenenti, i due superiori, alle vie ottiche e i due inferiori alle vie uditive (controllandone alcune attività riflesse). Superiormente il solco crociato, formato dai tubercoli, si espande appianandosi dando posto alla ghiandola endocrina epifisi.
Il midollo allungato e il ponte formano il romboencefalo. Il romboencefalo più il mesoencefalo fomano il tronco encefalico.
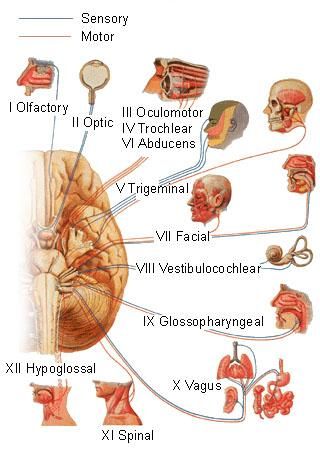 I nervi cranici (nervi encefalici) sono un gruppo di nervi che invece di avere origine dal midollo spinale, partono direttamente dal tronco encefalico costituendo l'innervazione motrice e sensitiva della testa nonchè l'innervazione viscerale, sia in uscita che in entrata (efferente e afferente), degli importanti centri del sistema nervoso parasimpatico che controllano l'attività dei visceri della testa, del collo, del torace e dell'addome. Nell'anatomia umana ci sono dodici paia di nervi cranici (destri e sinistri), numerati dall'alto verso il basso con numeri romani.
Essi hanno un'organizzazione più complessa rispetto ai nervi spinali. Anche i nuclei dei nervi cranici (nuclei encefalici), ossia le zone in cui sono situati i corpi cellulari dei neuroni corrispondenti, sono più complessi di quelli spinali. La sostanza grigia, a differenza del midollo spinale, non è raccolta in un'unica formazione ma è suddivisa in vari nuclei in parte ai nervi encefalici.
Quasi tutti i nervi cranici sono connessi a dei nuclei di materia grigia all'interno del tronco encefalico e gli assoni da e per i nervi cranici incontrano le loro sinapsi all'interno di questi nuclei.
Come nel midollo spinale, anche i nuclei del tronco encefalico sono divisi in anteriori motori e posteriori sensoriali.
Nel tronco encefalico vi sono inoltre nuclei non direttamente connessi all'origine dei nervi encefalici, detti nuclei propri (fra i quali citiamo, per l'importanza rivestita nel sistema dell'equilibrio e postura, i nuclei vestibolari e i nuclei oculomotori), e la formazione (sostanza) reticolare. La sostanza reticolare è un'antica formazione nervosa dalla citoarchitettonica caratteristica, costituita da varie tipologie neuronali aggregate in piccoli nuclei, nell'ambito di un complesso reticolato di fibre nervose che si estende per tutto il tronco encefalico. Le fibre della sostanza reticolare appartengono sia vie nervose ascendenti che discendenti influenzando tutto il sistema nervoso.
I nervi cranici (nervi encefalici) sono un gruppo di nervi che invece di avere origine dal midollo spinale, partono direttamente dal tronco encefalico costituendo l'innervazione motrice e sensitiva della testa nonchè l'innervazione viscerale, sia in uscita che in entrata (efferente e afferente), degli importanti centri del sistema nervoso parasimpatico che controllano l'attività dei visceri della testa, del collo, del torace e dell'addome. Nell'anatomia umana ci sono dodici paia di nervi cranici (destri e sinistri), numerati dall'alto verso il basso con numeri romani.
Essi hanno un'organizzazione più complessa rispetto ai nervi spinali. Anche i nuclei dei nervi cranici (nuclei encefalici), ossia le zone in cui sono situati i corpi cellulari dei neuroni corrispondenti, sono più complessi di quelli spinali. La sostanza grigia, a differenza del midollo spinale, non è raccolta in un'unica formazione ma è suddivisa in vari nuclei in parte ai nervi encefalici.
Quasi tutti i nervi cranici sono connessi a dei nuclei di materia grigia all'interno del tronco encefalico e gli assoni da e per i nervi cranici incontrano le loro sinapsi all'interno di questi nuclei.
Come nel midollo spinale, anche i nuclei del tronco encefalico sono divisi in anteriori motori e posteriori sensoriali.
Nel tronco encefalico vi sono inoltre nuclei non direttamente connessi all'origine dei nervi encefalici, detti nuclei propri (fra i quali citiamo, per l'importanza rivestita nel sistema dell'equilibrio e postura, i nuclei vestibolari e i nuclei oculomotori), e la formazione (sostanza) reticolare. La sostanza reticolare è un'antica formazione nervosa dalla citoarchitettonica caratteristica, costituita da varie tipologie neuronali aggregate in piccoli nuclei, nell'ambito di un complesso reticolato di fibre nervose che si estende per tutto il tronco encefalico. Le fibre della sostanza reticolare appartengono sia vie nervose ascendenti che discendenti influenzando tutto il sistema nervoso.
Il ponte presenta importanti connessioni col cervelletto (assieme costituiscono il metencefalo) che è la seconda regione dell'encefalo più voluminosa dopo il "cervello superiore". Esso si appoggia posteriormente al romboencefalo piegandone anteriormente l'asse, così che il ponte si trova ad appoggiarsi direttamente sulla base cranica.
Il cervelletto è diviso in due emisferi cerebellari divisi dal verme, sulla cui faccia anteriore è presente il flocculo.
Esso è costituito da regioni antiche (verme, flocculo e il quarto anteriore dei due emisferi) e regioni recenti (compartimenti posteriori degli emisferi), che rappresentano la parte più estesa, con funzioni diverse.
All'interno della sostanza bianca, in profondità, sono situati i nuclei cerebellari (ammassi di sostanza grigia) di cui, il nucleo dentato, risulta particolarmente sviluppato. La sostanza grigia della corteccia cerebellare (ca. 1 mm di spessore) si ripiega in superficie per accogliere il maggior numero possibile di neuroni.
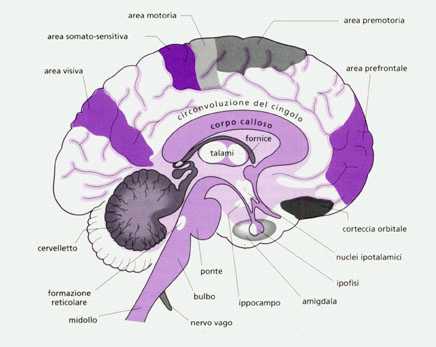
Superiormente al mesoencefalo (mesencefalo) troviano il diencefalo composto fondamentalmente dal talamo destro e sinistro e dall'ipotalamo posto al centro, quest'ultimo è collegato con la ghiandola endocrina ipofisi (sistema neuroendocrino), con la sostanza reticolare e col sistema limbico. Il sistema o lobo limbico rappresenta quei giri corticali, filogeneticamente più antichi, che circondano ad anello il tronco encefalico e comprende l'amigdala, l'ippocampo, i corpi mammillari ed il giro del cingolo o cingolato (immagine). Il sistema limbico è un circuito costituito da un insieme di centri corticali e subcorticali fra loro interconnessi da proiezioni di fasci di fibre nervose (tra cui le più importanti sono la fornice che unisce ippocampo e diencefalo e la stria terminale che collega l'amigdala all'ippocampo) che trasportano specifici neurotrasmettitori. Notevole è l'importanza dell'insieme ipotalamo-sistema limbico relativamente a numerose funzioni vitali dell'organismo.
Il telecenfalo, considerato dai fisiologici e dai neuroclinici il "cervello" in senso stretto, è la parte di gran lungo più voluminosa dell'encefalo e ricopre tutte le precedenti strutture.
Il telencefalo risulta diviso in due emisferi (destro e sinistro) uniti tra loro profondamente, all'interno della scissura interemisferica (in cui penetrano le meningi), da fasci di fibre nervose che formano un nucleo di sostanza bianca chiamato corpo calloso.
Ogni emisfero è diviso in 4 lobi (frontale, parietale, temporale e occipitale) in cui si identificano aree (aree di proiezione) con funzioni motorie o sensitive prevalentemente specifiche.
La parete degli emisferi cerebrali è la corteccia cerebrale costituita da sostanza grigia ricca di neuroni che riveste la sostanza bianca formata da fibre nervose (assoni).
Per far fronte all'aumento del numero delle cellule cerebrali nel corso dello sviluppo evolutivo (nell'uomo sono ca. 10 miliardi), la corteccia cerebrale, come nel cervelletto,
ha dovuto ripiegarsi su se stesso, formando le circonvoluzioni (nel regno animale è possibile dimostrare che all'aumentare delle capacità intellettive corrisponde un aumento della complessità delle circonvoluzioni cerebrali).
L'organizzazione dell'encefalo non si differenzia in modo sostanziale da quella del midollo spinale ma solo nel telencefalo e nel cervelletto è la sostanza grigia (corpi di cellule nervose e fibre nervose amieliniche) a circondare la bianca (fibre nervose mieliniche).
Il vantaggio di questo tipo di organizzazione, rispetto a quallo presente nel midollo spinale e nel troncoencefalo (sostanza bianca all'esperno e grigia all'interno), è di consentire vie di collegamenti più brevi (questo vantaggio è sfruttato anche nei computer moderni in cui le chips sono poste esternamente e il cablaggio internamente).
Analogamente al cervelletto, all'interno della sostanza bianca, in profondità, sono situati i nuclei della base (globo pallido, sostanza nera, putamen e nucleo caudato; quest'ultimi due spesso compresi nel "corpo striato").
Quanto più una porzione cerebrale è recente, tanto più complesse sono le sue funzioni. Il midollo allungato contiene i centri vegetativi vitali (in particolare repiratori e circolatori) e collabora
strettamente con il ponte, il cervelletto e il mesencefalo in quella che sembra essere la loro funzione principale ovvero la regolazione dell postura in statica e in movimento (situazione simile è presente nei vertebrati inferiori).
La formazione reticolare disseminata nel tronco encefalico funge da sistema relativamente autonomo capace di influenzare le strutture nervose di tutto l’asse cerebrospinale: attraverso vie nervose discendenti regola funzioni somatiche e vegetative quali tono posturale, attività motoria, respiro, pressione arteriosa, frequenza cardiaca, tramite le vie nervose ascendenti, le quali raggiungono diencefalo e corteccia cerebrale, modula il flusso degli impulsi provenienti dai recettori sensoriali e provvede ad attivare la corteccia cerebrale mantenendo lo stato di attenzione e veglia.
Il talamo è il centro riflesso delle emozioni che provocano il pianto e il riso controllandone la motilità relativa e la sensibilità degli imput ricevuti dai recettori periferici.
L'ipotalamo e il sistema limbico, con il quale è funzionalmente e strutturalmente collegato, presiedono a quei meccanismi vitali che hanno lo scopo di mantenere costanti le condizioni dell'ambiente interno (omeostasi) e di provvedere alla conservazione dell'individuo e della specie, esercitando il controllo sulle sue emozioni e sulla sessualità: regolazione del sistema nervoso autonomo e dell'apparato endocrino, della temperatura corporea, del ciclo sonno/veglia, della frequenza cardiaca, della pressiona arteriosa, dell'osmolarità del sangue, dell'assunzione di cibo e acqua, della secrezione, acida dello stomaco, del metabolismo dei glicidi e dei grassi, delle emozioni e delle funzioni sessuali.
L'ippocampo e' inoltre importante per l'apprendimento e la memoria. La caratteristica peculiare dell'ipotalamo è quella di ricevere degli imput direttamente dall'esterno del cervello. L'ipotalamo infatti riceve direttamente il segnale luce-buio (tratto retino-ipotalamico) e quello olfattivo (via olfattiva basale), in più, esso costituisce, tramite un fascio di fibre nervose e una rete di vasi sanguinei (eminenza mediana dell'ipotalamo) che, come accade anche per gli altri organi circumventricolari, non presentano la barriera emato-encefalica, la parte posteriore della ghiandola endocrina ipofisi, detta neuroipofisi.
La neuroipofisi è così formata dai prolungamenti assonici di neuroni il cui corpo cellulare è sito nei nuclei sovraottico e paraventricolare dell’ipotalamo. La neuroipofisi risulta quindi essere in realtà non una ghiandola ma un organo nervoso a tutti gli effetti, un'estroflessione cerebrale formata da tessuto nervoso. Più precisamente la neuropifisi è un organo "neuroemale" in quanto i neurotrasmettitori secreti a livello delle sue sinapsi terminali invece di stimolare il neurone successivo (come accade nelle sinapsi propriamente dette), si riversano nel sangue generando una risposta più generalizzata. Questo collegamento diretto sistema nervoso centrale-apparato endocrino (sistema neuroendocrino) assume grande rilievo nella regolazione generale dei processi vitali. L'ipotalamo inoltre è esso stesso anche una ghiandola endocrina i cui ormoni, agenti sull'adenoiposi (fattori ipotalamici), creano con essa un doppio legame: strutturale (neuroipofisi) e umorale, tramite ormoni (adenoipofisi). Ulteriori ormoni secreti dalla neuroipofisi sono l’ADH (ormone antidiuretico) e l’ossitocina.
L'ipotalamo grazie alle sue connessioni con la neocorteccia, col sistema limbico, con la sostanza reticolare, col sistema nervoso vegetativo e col sistema endocrino è considerato da molti studiosi la struttura limite tra somatico e psichico, quella cioè in grado di commutare il segnale pschico in chimico e viceversa. E' grazie all'ipotalamo che gli aspetti mentali, emotivi e istintivi trovano espressione nel soma.
Nel telencefalo, infine, hanno sede le funzioni cerebrali "superiori" quali il pensiero, il linguaggio, la programmazione motoria ecc.
 Il canale formato dalle vertebre costituisce un'importante protezione per il delicato midollo spinale (che insieme all'encefalo costituisce il sistema nervoso centrale).
Racchiuso nel sacco durale meningeo, Il midollo spinale ha forma cilidrica, con diametro medio 8-10 mm, e si estende dal grande forame occipitale (base cranio, corpo della prima vertebra cervicale o atlante), continuandosi dal midollo allungato, a circa la prima o seconda vertebra lombare (non occupa quindi il canale vertebrale per tutta la sua lunghezza) anche se a livello sacrale è presente il midollo sacrale con struttura simile. Qui il midollo spinale termina a forma di cono (cono midollare) e prosegue caudalmente, quale sottile struttura fibrovascolare circondata dai nervi della cauda equina (filo o filum terminale), ancorandosi al coccige (oltre che dal filum terminale, il midollo spinale è mantenuto in posizione fisiologica dai legamenti dentati che lo connettono bilateralmente al sacco durale).
Il canale formato dalle vertebre costituisce un'importante protezione per il delicato midollo spinale (che insieme all'encefalo costituisce il sistema nervoso centrale).
Racchiuso nel sacco durale meningeo, Il midollo spinale ha forma cilidrica, con diametro medio 8-10 mm, e si estende dal grande forame occipitale (base cranio, corpo della prima vertebra cervicale o atlante), continuandosi dal midollo allungato, a circa la prima o seconda vertebra lombare (non occupa quindi il canale vertebrale per tutta la sua lunghezza) anche se a livello sacrale è presente il midollo sacrale con struttura simile. Qui il midollo spinale termina a forma di cono (cono midollare) e prosegue caudalmente, quale sottile struttura fibrovascolare circondata dai nervi della cauda equina (filo o filum terminale), ancorandosi al coccige (oltre che dal filum terminale, il midollo spinale è mantenuto in posizione fisiologica dai legamenti dentati che lo connettono bilateralmente al sacco durale).
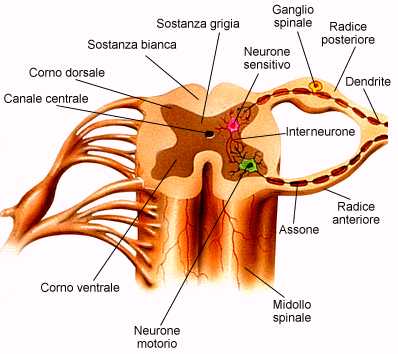
Al pari della colonna vertebrale, il midollo spinale è diviso in segmenti (segmenti midollari). Sezionando trasversalmente uno di questi segmenti, si trova una disposizione opposta a quella di cervello e cervelletto e uguale a quella del tronco encefalico. Infatti,
nel midollo spinale la sostanza grigia è all'interno e la sostanza bianca all'esterno.
La sostanza grigia del midollo spinale, composta soprattutto dai corpi delle cellule nervose (neuroni), ha una forma che ricorda una farfalla. Questi ammassi di corpi cellulari rappresentano i nuclei spinali.
Nelle ali anteriori, dette corna anteriori, sono presenti, tra l'altro, i corpi cellulari dei motoneuroni alfa e gamma (classificazione delle fibre nervose), che innervano la muscolatura scheletrica. Gli assoni di questi neuroni fuoriescono dal midollo spinale, avvolti in una spessa guaina mielinica bianca, tramite le radici anteriori.
La radice anteriore è pertanto motoria ossia contiene fibre motrici somatiche (volontarie) ed effettrici viscerali. A livello delle corna anteriori arrivano i segnali motori derivanti dalla corteccia cerebrale e dal tronco encefalico (nuclei vestibolari).
Le ali posteriori, corna posteriori, si estendono fino all'imbocco delle radici posteriori, dove si raccolgono, in un ganglio spinale, le fibre sensitive (sensibilità estero-propriocettiva). La radice posteriore è quindi sensitiva e contiene fibre afferenti, somatiche e viscerali. Qui terminano anche gli assoni deputati alla soppressione del dolore provenienti dal tronco encefalico.
Secondo la teoria del "gate control" il neurotrasmettitore delle fibre del tatto, encefalina, inibisce la trasmissione sinaptica bloccando la percezione del dolore; stimoli emotivi intensi (shock) e stimoli tattili (digitopressione, agopuntura) possono così rendere temporaneamente insensibili al dolore.
I corni laterali (nucleo intermedio laterale) sono piccole sporgenze a metà strada tra corna anteriore e posteriore. Le loro cellule, tramite le radici anteriori, inviano assoni agli organi innervati dal sistema nervoso vegetativo. Da questa colonna grigia laterale, nascono alcune fibre di natura ortosimpatica (nel tratto toracico del midollo spinale), e parasimpatica (a livello del midollo sacrale).
Al centro della sostanza grigia vi è un sottile canale centrale (rivestito da epitelio ependimale e in cui può essere presente liquor cerebrospinale) che è un residuo embrionale e non di rado è occluso o dilatato in cisti.
Immediatamente adiacenti alla sostanza grigia decorrono brevi fasci di fibre nervose, detti fasci fondamentali, che collegano 4-5 segmenti midollari confinanti circondando, come un sottile mantello, l'intera sostanza grigia. Essi provvedono a schemi di riflessi spinali complessi ovvero che non riguardano la muscolatura di un unico segmento (riflessi polisegmentali); tramite i riflessi spinali, ogni singolo segmento mdollare ha la capacità di controllare autonomamente funzioni motorie specifiche che richiedono rapidità di risposta.
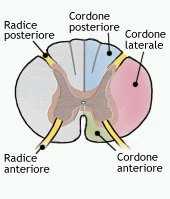 La massa della sostanza bianca è invece formata da lunghi fasci midollari ascendenti e discendenti che collegano encefalo e midollo spinale. Questi fasci decorrono in tre cordoni a destra e tre a sinistra, separati fra loro dalla fessura centrale midollare, dal corno posteriore e da quello anteriore. L'ordine per una contrazione muscolare di precisione (ad esempio riguardanti movimenti delle dita della mano) viaggia lungo fibre che provengono dall'area motoria primaria encefalica attraverso la via piramidale ossia tramite il fascio del midollo spinale (fascio corticospinale) e lungo il fascio corticonucleare diretto ai nuclei dei nervi cranici del tronco encefalico. Il 90% delle fibre del fascio piramidale si incrocia nel midollo allungato, passando dal lato opposto (decussazione), e discende nella compagine del fascio corticospinale laterale. Il restante 10% non si incrocia nel midollo allungato e decorre nel fascio corticospinale anteriore, incrociandosi immediatamente prima dell' entrata nella sostanza grigia delle corna anteriori. Le fibre di entrambi i fasci terminano direttamente (meno di 1/3) oppure tramite un interneurone sulle cellule motorie delle corna anteriori, formando una sinapsi col motoneurone alfa e attivando in tal modo un riflesso spinale.
La massa della sostanza bianca è invece formata da lunghi fasci midollari ascendenti e discendenti che collegano encefalo e midollo spinale. Questi fasci decorrono in tre cordoni a destra e tre a sinistra, separati fra loro dalla fessura centrale midollare, dal corno posteriore e da quello anteriore. L'ordine per una contrazione muscolare di precisione (ad esempio riguardanti movimenti delle dita della mano) viaggia lungo fibre che provengono dall'area motoria primaria encefalica attraverso la via piramidale ossia tramite il fascio del midollo spinale (fascio corticospinale) e lungo il fascio corticonucleare diretto ai nuclei dei nervi cranici del tronco encefalico. Il 90% delle fibre del fascio piramidale si incrocia nel midollo allungato, passando dal lato opposto (decussazione), e discende nella compagine del fascio corticospinale laterale. Il restante 10% non si incrocia nel midollo allungato e decorre nel fascio corticospinale anteriore, incrociandosi immediatamente prima dell' entrata nella sostanza grigia delle corna anteriori. Le fibre di entrambi i fasci terminano direttamente (meno di 1/3) oppure tramite un interneurone sulle cellule motorie delle corna anteriori, formando una sinapsi col motoneurone alfa e attivando in tal modo un riflesso spinale.
Encefalo e midollo spinale formano il sistema nervoso centrale.
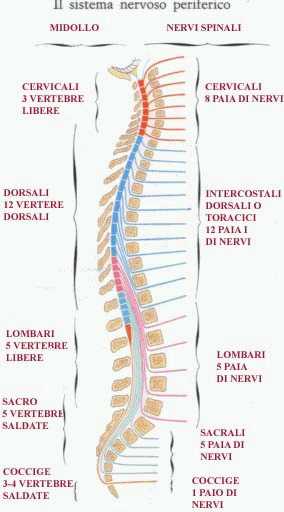 Ciascun segmento midollare emette, uno a destra e uno a sinistra, attraverso il foro di coniugazione o intervertebrale formato da due vertebre adiacenti, i nervi spinali che presentano la confluenza di una radice anteriore (ventrale), da cui partono fibre efferenti (motorie somatiche ed effettrici viscerali) con destinazione l'organo effettore, ghiandola o muscolo, e una posteriore (dorsale), a cui giungono fibre afferenti sensitive (somatiche e viscerali), derivanti dai recettori sensoriali. Tutti i nervi spinali quindi risultano misti ossia composti da una componente motoria e una sensitiva.
Ciascun segmento midollare emette, uno a destra e uno a sinistra, attraverso il foro di coniugazione o intervertebrale formato da due vertebre adiacenti, i nervi spinali che presentano la confluenza di una radice anteriore (ventrale), da cui partono fibre efferenti (motorie somatiche ed effettrici viscerali) con destinazione l'organo effettore, ghiandola o muscolo, e una posteriore (dorsale), a cui giungono fibre afferenti sensitive (somatiche e viscerali), derivanti dai recettori sensoriali. Tutti i nervi spinali quindi risultano misti ossia composti da una componente motoria e una sensitiva.
Appena fuori dal foro di coniugazione ogni nervo spinale si divide in 2 rami (entrambi sempre sia motori che sensitivi).
Il ramo posteriore (dorsale) non presenta fibre visceromotrici e innerva, dal punto di vista motorio, la muscolatura spinale e, dal punto di vista sensitivo, la regione cutanea del dorso.
Il ramo anteriore (ventrale) del nervo spinale costituisce la porzione più voluminosa e funzionalmente importante. Esso possiede fibre somatomotrici e visceromotrici, somatosensitive e viscerosensitive e da esso partono due diramazioni (dx e sx), o rami comunicanti (fibre simpatiche pregangliari), che terminano in un ganglio simpatico per poi interessare i relativi organi bersaglio.
I rami anteriori dei nervi toracici percorrono isolatamente i rispettivi spazi intercostali, quali nervi intercostali mentre gli altri rami anteriori si intrecciano e anastomizzano tra loro formando 6 plessi nervosi: plesso nervoso cervicale, brachiale, lombare, sacrale, coccigeo, pudendo.
Le fibre somatiche motrici e sensitive del ramo anteriore innervano muscoli e pelle degli arti e della superficie antero-laterale del corpo.
Tutte le parti del corpo, a eccezione del viso, del tratto gastrointestinale e di parti della muscolatura cervicale (zone innervate dai nervi cranici), sono innervate dai nervi spinali somatici (volontari).
A differenza della colonna vertebrale, il midollo spinale è dotato di 8 segmenti cervicali (invece di 7). I 12 segmenti toracici, 5 lombari, 5 sacrali e 1-2 coccigei sono invece in numero corrispondente alle relative vertebre. I nervi spinali cervicali fuoriescono dal foro di coniugazione superiore alla vertebra corrispondente, fatta eccezione dei due dell'VIII segmento (C8 dx e C8 sn) che escono da quello inferiore, così come accade per tutti i restanti segmenti. Affinchè ciò accada, essendo il midollo spinale più corto rispetto al canale vertebrale (a partire dal quarto mese di vita fetale il rachide si sviluppa più rapidamente del midollo spinale), le radici nervose dei segmenti lombari, sacrali e coccigei divengono sempre più distese verso il basso, formando così un fascio di sottili fibre nervose pressochè parallele che ricordano la coda di un cavallo (da cui la denominazione di cauda equina).
I nervi spinali e i nervi cranici costituiscono i nervi encefalospinali. Essi insieme ai relativi gangli formano il sistema nervoso periferico.
ll sistema nervoso periferico è composto da tutti i nervi (cranici e spinali) e i relativi gangli presenti all’esterno del sistema nervoso centrale (SNC). Il sistema nervoso periferico trasporta, tramite le fibre nervose afferenti, l’informazione sensoriale (estero-propriocettiva) al SNC, e, per mezzo delle fibre nervose efferenti (effettrici, motrici), i comandi dal SNC agli organi e tessuti. I corpi delle cellule nervose sono raggruppati nei gangli nervosi del sistema nervoso periferico e nei nuclei del midollo spinale e dell'encefalo, che costituiscono dei pool neuronali con specifiche caratteristiche di organizzazione ed elaborazione dei segnali.
Il sistema nervoso periferico viene suddiviso in sistema nervoso somatico (o volontario) e il sistema nervoso autonomo (o vegetativo o viscerale o involontario). Entrambi vengono controllati e coordinati dal sistema nervoso centrale. La classica netta separazione tra il sistema nervoso vegetativo, considerato come controllore delle funzioni viscerali in maniera indipendente dalla nostra volontà, e il sistema nervoso volontario, controllato dalla ns. volontà, è oggi sempre più messa in discussione.
Il sistema nervoso periferico volontario o somatico consiste nella componente del sistema nervoso periferico che controlla i muscoli scheletrici (muscoli definiti volontari) e quindi principalmente i gesti motori. I corpi cellulari dei motoneuroni del sistema nervoso somatico si trovano nella sostanza grigia del midollo spinale e gli assoni raggiungono direttamente i muscoli scheletrici, con i quali si connettono tramite speciali sinapsi (placche motrici). Diverso è invece il discorso per le vie afferenti.
 Il sistema nervoso periferico vegetativo (autonomo, involontario, viscerale) - SNA rappresenta la porzione del sistema nervoso periferico che innerva le ghiandole, la muscolatura cardiaca e quella liscia di organi e vasi sanguigni, interessando, in tal modo, le funzioni del corpo normalmente sotto controllo inconscio (ritmo cardiaco e circolazione sanguigna, respirazione, digestione ecc.). Il sistema nervoso vegetativo (autonomo) è composto sia da fibre nervose efferenti (motrici) che afferenti (sensoriali); il 75% delle fibre del nervo vago sono afferenti. Nel sistema nervoso autonomo la via efferente è sempre costituita da due neuroni (a differenza di quella del sistema nervoso volontario che è costituita da un solo motoneurone): un neurone pregangliare col corpo cellulare posto nel sistema nervoso centrale e un neurone postgangliare, con il corpo cellulare al di fuori di esso, in un ganglio o nella parete dell'organo innervato.
Il sistema nervoso periferico vegetativo (autonomo, involontario, viscerale) - SNA rappresenta la porzione del sistema nervoso periferico che innerva le ghiandole, la muscolatura cardiaca e quella liscia di organi e vasi sanguigni, interessando, in tal modo, le funzioni del corpo normalmente sotto controllo inconscio (ritmo cardiaco e circolazione sanguigna, respirazione, digestione ecc.). Il sistema nervoso vegetativo (autonomo) è composto sia da fibre nervose efferenti (motrici) che afferenti (sensoriali); il 75% delle fibre del nervo vago sono afferenti. Nel sistema nervoso autonomo la via efferente è sempre costituita da due neuroni (a differenza di quella del sistema nervoso volontario che è costituita da un solo motoneurone): un neurone pregangliare col corpo cellulare posto nel sistema nervoso centrale e un neurone postgangliare, con il corpo cellulare al di fuori di esso, in un ganglio o nella parete dell'organo innervato.
Il sistema nervoso vegetativo si divide in simpatico o ortosimpatico (toraco-lombare), parasimpatico (cranio-sacrale e metasimpatico (enterico).
Il sistema nervoso ortosimpatico (simpatico o toraco-lombare) (parte del sistema nervoso vegetativo o autonomo, a sua volta rientrante nel sistema nervoso periferico) è composto da un'unità centrale di controllo, l'ipotalamo e dai relativi corpi neuronali, fibre nervose (efferenti e afferenti) e dai relativi gangli. Il sistema nervoso simpatico presenta coppie di neuroni efferenti (motrici, effettori) posti consecutivamente (I e II neurone). Il primo neurone effettore (pregangliare), localizzato nel corno laterale dei segmenti spinali che vanno dalla prima vertebra toracica (T1) alla seconda lombare (L2), emette un assone che si diparte, attraverso la radice anteriore del midollo spinale, insieme al nervo spinale (ramo anteriore), da cui si divide appena fuori dal foro di coniugazione spinale (foro intervertebrale) dirigendosi, come breve ramo comunicante bianco (fibre pregangliari mieliniche), al ganglio della catena dell'ortosimpatico del segmento spinale corrispondente (ganglio laterovertebrale o paravertebrale) o, quali nervi splancnici (viscerali), ai gangli prevertebrali (ad es. mesenterici e celiaco) posti frontalmente alla colonna vertebrale. I gangli paravertebrali formano le catene (o tronchi) dell'ortosimpatico che appaiono come due fili di perle poste parallelamente, una destra e l'altra a sinistra, lungo tutta la colonna vertertebrale. Tali gangli (le "perle") sono connessi fra di loro da sottili fasci nervosi e ciò consente a una fibra pregangliare di prendere contatti anche con altri segmenti, sia superiori che inferiori. A livello dei gangli paravertebrali e prevertebrali, le fibre pregangliari contraggono sinapsi con i corpi cellulari del II neurone (neurone postgangliare); da qui quindi partono le fibre nervose amieliniche (assoni) postgangliari che raggiungono gli organi bersaglio seguendo i vasi arteriosi vicini (rami vascolari) o vanno distalmente tornando al nervo spinale (rami comunicanti grigi).
Diversamente da ciò che accade di norma, la porzione midollare delle surrenali (due piccole ma importantissime ghiandole endocrine poste ognuna sopra il polo superiore dei reni), oltre a presentare una componente nervosa esclusivamente simpatica, viene innervata, attraverso i nervi splancnici (viscerali), da fibre nervose pregangliari del simpatico che non si interrompono, ovvero che non formano sinapsi con i gangli della catena paravertebrale o prevertebrali dell'ortosimpatico, connettondosi direttamente a un determinato gruppo di cellule della midollare, cellule cromaffini (formando in sostanza un ganglio interno), stimolandole a produrre le catecolamine adrenalina, noradrenalina e, in minima quantità, dopamina. Ci troviamo qui di fronte a una via direttissima che collega il cervello, tramite il sistema nervoso simpatico, alle surrenali. La visione del pericolo, infatti, mette in funzione questa via immediata (fase di allarme della reazione di stress), inducendo a produrre, da parte delle cellule della midollare del surrene, una miscela “allarmante” composta per l’80% di adrenalina e il 20% di noradrenalina, in quantità dieci volte maggiore del normale. Se però il pericolo perdura (fase di resistenza della reazione di stress), allora entreranno in scena ulteriori significativi cambiamenti (attivazione asse HPA) che coinvolgeranno ancora una volta le surrenali (questa volta la zona corticale), organi primari della reazione di stress.
Gli effetti del sistema nervoso ortosimpatico sono facilmente comprensibili se interpretati in relazione alla "reazione di attacco o difesa" (stress fisico e psichico): aumentano della frequenza cardiaca, della pressione arteriosa, della ventilazione (con dilatazione dei bronchi), del tono dei muscoli scheletrici, dilatazione delle arterie coronariche e dei vasi dei muscoli scheletrici, contrazione dei vasi sanguigni periferici (cute) e degli organi viscerali (tranne cuore e polmoni), dilatazione delle pupille e accomodamento per la visione lontana (tramite rilassamento del muscolo ciliare), rallentamento dei processi digestivi e dell'attività del sistema immunitario, "annebbiamento" dell'attività cognitiva ecc. (schema reazione di stress). ll sistema nervoso parasimpatico è suo antagonista contrastandone e bilanciandone gli effetti.
Le fibre pregangliari ortosimpatiche utilizzano come neurotrasmettitore l'acetilcolina e il peptide vasointestinale (VIP), quelle postgangliari la noradrenalina, l'adrenalina, la dopamina (prodotte dalla midollare del surrene sotto lo stimolo dall'acetilcolina) e il neuropeptide Y (NPY); il sistema nervoso simpatico è noto anche come sistema adrenergico.
Come il sistema simpatico, anche il sistema nervoso parasimpatico (o cranio-sacrale) rientra nel sistema nervoso vegetativo o autonomo del sistema nervoso periferico ed è regolato dall'ipotalamo. Anatomicamente è situate nelle zone del canale vertebrale non occupate dall'ortosimpatico ossia il tronco encefalico e il midollo sacrale. Il midollo sacrale, come il midollo spinale propriamente detto, è racchiuso nel sacco durale (nel filum terminale) e presenta la sostanza grigia con la classica struttura a farfalla) e rappresenta la sede delle attività riflesse parasimpatiche per la minzione, l'erezione e la motilità anorettale; l'intervento dei centri sovraspinali consente il controllo volontario di tali attività riflesse. L'innervazione parasimpatica pertanto decorre lungo i nervi cranici, di cui il X o nervo vago innerva gli organi della cavità toracica e addominale (quasi esclusivamente dipendenti da questo nervo) mentre i restanti raggiungono gli organi cranici, e lungo gli assoni dei neuroni parasimpatici presenti nei corni laterali dei segmenti spinali che vanno dalla seconda alla quarta vertebra sacrale (S2-S4). Questi ultimi decorrono poi quasi tutti nel nervo pudendo innervando organi genitali, vescica e intestino retto.
Tutti i nervi parasimpatici contengono lunghe fibre pregangliari (che utilizzano come neurotrasmettitori l'acetilcolina e il peptide vasointestinale VIP) in quanto, a differenza del sistema nervoso ortosimpatico, i gangli parasimpatici sono situati in prossimità o all'interno degli organi innervati.
Sotto il dominio del sistema parasimpatico dimorano il rilassamento, il riposo, la rigenerazione, la digestione (aumento delle secrezioni digestive, della peristalsi ecc.), l'immagazzinamento di energia ecc. ("rest and digest"). Pertanto l'azione del parasimpatico si contrappone a quella del simpatico, il più delle volte, all'interno di un raffinato bilanciamento avente come obiettivo l'omeostasi.
La maggior parte degli organi è innervata da entrambe le componenti del sistema nervoso vegetativo (ortosimpatica e parasimpatica). Vi sono però alcune parti del corpo che ricevono un’innervazione esclusivamente simpatica. Tra queste vi è il fegato e la porzione midollare delle surrenali (due piccole ma importantissime ghiandole endocrine poste ognuna sopra il polo superiore dei reni).
Oltre a presentare nervi misti (il nervo vago è composto per il 75% da fibre nervose sensoriali), il sistema nervoso vegetativo invia al midollo spinale sensazioni quali, per esempio, dolore, stato di riempimento dello stomaco e della vescica ecc. Tali fibre nervose arrivano normalmente alla radice posteriore del midollo spinale, tramite le vie afferenti, non distinguendosi però in ortosimpatiche o parasimpatiche. Pertanto, poichè nei segmenti del midollo spinale convergono afferenze della sensibilità dolorifica degli organi interni e afferenze cutanee, il dolore viscerale può essere proiettato e quindi percepito sulle aree cutanee, dermatomeri, del relativo segmento midollare (cuore-faccia interno braccio sinistro, cistifellea-addome superiore destro, pancreas-addome sinistro ecc.).
Descrivendo il sistema nervoso autonomo (vegetativo) occorre ricordare l'esistenza di una terza componente vegetativa: il sistema nervoso metasimpatico (enterico), importante quanto le altre due (ortosimpatica e parasimpatica). Il sistema nervoso metasimpatico è presente all'interno delle pareti dell'intestino e dello stomaco come rete nervosa formata da circa cento milioni di neuroni, collegata alle fibre nervose ortosimpatiche e parasimpatiche (plesso sottomucoso di Meissner e motorio mienterico di Auerbach). Tale sistema nervoso enterico innerva il tratto gastro-intestinale, la colecisti e il pancreas. Ricerche recenti hanno potuto affermare che questa rete svolge un ruolo in gran parte indipendente dal sistema nervoso centrale (SNC). Data l'importanza attruibuita dai fisiologi a tale struttura, essa viene definita come "cervello addominale o "cervello enterico". E' chiaro, data la grande attiguità, che l'alimentazione avrà sul sistema metasimpatico una grossa influenza.
In conclusione, oggi, grazie all'avvento della psiconeuroimmunologia, sappiamo che il sistema nervoso costituisce una fittissima rete di comunicazione (solo nel cervello vi sono 10 miliardi di neuroni) diffusa in tutto il corpo e strettamente interconnessa con due altri importanti sistemi di regolazione dell'organismo: il sistema endocrino e il sistema immunitario. Al contempo, anatomia e fisiologia del sistema nervoso dipendono in modo importante dal sistema connettivo.
Nel neurone la trasmissione del segnale lungo l'assone e il dendrite è di tipo elettrico (grazie a una variazione della polarizzazione della membrana) mentre a livello sinaptico è di tipo chimico. I neurotrasmettitori (neuromediatori) sono molecole che consentono la trasmissione degli impulsi nervosi fra due neuroni attraverso le sinapsi o tra fibra nervosa terminale e organo effettore. Il neurotrasmettitore, all'arrivo dell'impulso nervoso (potenziale d'azione), viene liberato dalle vescicole presinaptiche, migra nello spazio intersinaptico fino a fissarsi su specifici recettori della membrana postsinaptica causando la depolarizzazione della membrana e quindi la trasmissione dell'impulso. Nel giro di pochi millisecondi viene disattivato, tramite specifici enzimi (colinesterasi, monoaminossidasi ecc.), e riassorbito nelle membrane presinaptiche (reuptake).
Esistono numerosi neurotrasmettitori con effetto eccitatorio e inibitorio. L'azione di uno specifico neurotrasmettitore dipende principalmente dalle caretteristiche del recettore postsinaptico (uno stesso neurotrasmettitore può avere effetti inibitori o eccitatori in base al recettore a cui si lega). Oggi sappiamo che un neurone è in grado di sintetizzare diversi neurotrasmettitori nelle diversi sinapsi che contrae, anche se utilizza sempre lo stesso nella medesima sinapsi.
I principali neurotrasmettitori sono: acetilcolina (presente nei neuroni di tipo colinergico quali i motoneuroni e neuroni effettori viscerali pregangliari), adrenalina, noradrenalina e dopamina (catecolamine secrete dalla midollare del surrene e dalle terminazioni nervose postgangliari del sistema nervoso orosimpatico), serotonina (con azione essenzialmente inibitoria, interviene, fra l'altro, nei ritimi sonno-veglia, nel controllo del dolore e delle funzioni ipotalamiche), aminoacidi GABA (acido-gamma-amino-butirrico, principale neurotrasmettitore inibitorio con presenza ubiquitaria), aspartato e glutammato (principali neurotrasmettitori eccitatori del sistema nervoso centrale).
Oltre a questi vi sono molecole più complesse (vere e proprie proteine), i neuropeptidi, che possono fungere da neurotrasmettitori: glicina (neurotrasmettitore inibitorio, analogo al GABA, presente negli interneuroni del midollo spinale), peptide vasointestinale (VIP), neuropeptide Y (NPY), sostanza P, somatostatina, bombesina, istamina, neurotensina, encefalina e prostaglandina che, nel sistema nervoso centrale, possono fungere sia da neurotrasmettitori sia da neurormoni agendo a distanza su bersagli di natura endocrina. Talvolta i neuropeptidi possono agire da modulatori sinaptici (venendo secreti assieme ai neurotrasmettitori a livello sinaptico) organizzando i segnali specifici ed elementari dei normali neurotrasmettitori in un comportamento complesso.
I neurotrasmettitori hanno un ruolo essenziale, oltre che per la trasmissione degli impulsi nervosi, nella funzione di regolazione-modulazione dell'attività globale del sistema nervoso centrale, incluse le attività cognitive ed emotive.
In linea generale, la velocità di conduzione dell'impulso è tanto più elevata quanto maggiore è il diamentro della fibra nervosa ed esso va da 0,2 a 20 µm (1 µm = 1 micrometro, corrisponde a 1 milionesimo di metro), con velocità di conduzione da 0,5 a 120 metri/secondo. La vlocità di conduzione è inoltre aumentata in presenza della guaina mileinica.
Esistono due tipi di classificazione delle fibre nervose una classificazione generale, ossia che riguarda tutti i tipi di fibre, e una specifica per le fibre sensitive.
La classificazione generale (classificazione di Erlanger e Gasser) risulta:
- Fibre di tipo o gruppo A, sono le tipiche fibre mieliniche dei nervi spinali e sono a loro volta suddivise in alfa, beta, gamma, delta, con diametro e quindi velocità di conduzione decrescente (velocità di conduzione da 120 a circa 6 m/s, diametro da 20 a 1 µm);
- Fibre di tipo o gruppo B, sono le fibre pregangliari (mieliniche) del sistema nervoso autonomo, con valori di velocità di conduzione e diametro fra il tipo A e C.
- Fibre di tipo o gruppo C, fibre amieliniche di piccolo diametro e quindi a bassa velocità di conduzione, che costituiscono più della metà delle fibre sensitive dei nervi periferici e la totalità delle fibre postgangliari del sistema nervoso autonomo (velocità di conduzione da 2 a 0,5 m/s, diametro inferiore a 1,2 µm).
- Fibre tipo Ia, provenienti dalle terminazioni anulospirali dei fusi neuromuscolari, corrispondono al tipo A alfa della classificazione generale (diametro medio 17 µm);
- Fibre tipo o gruppo Ib, derivano dagli organi muscolo-tendinei del Golgi, corrispondono anch'esse al tipo A alfa (diametro medio 16 µm);
- Fibre tipo o gruppo II, provenienti dalla maggior parte dei meccanorecettori per la sensibilità cutanea fine (esterocettori cutanei), e dalle terminazioni secondarie e a fiorame dei fusi neuromuscolari, corrispondono alle fibre A beta e gamma della classificazione generale (circa 8 µm di diametro);
- Fibre tipo o gruppo III, relative ai recettori intersiziali, responsabili della sensibilità tattile grossolana, della termica e delle sensazioni dolorifiche puntiformi, corrispondono alle fibre A delta della classifica generale (circa 3 µm di diametro);
- Fibre tipo o gruppo IV, riguardano anch'esee i recettori intersiziali, amieliniche, trasportano impulsi della sensibilità dolorifica, termica, del prurito, tattile grossolana, appartengono al tipo C della classificazione generale (diametro 0,5-2 µm).
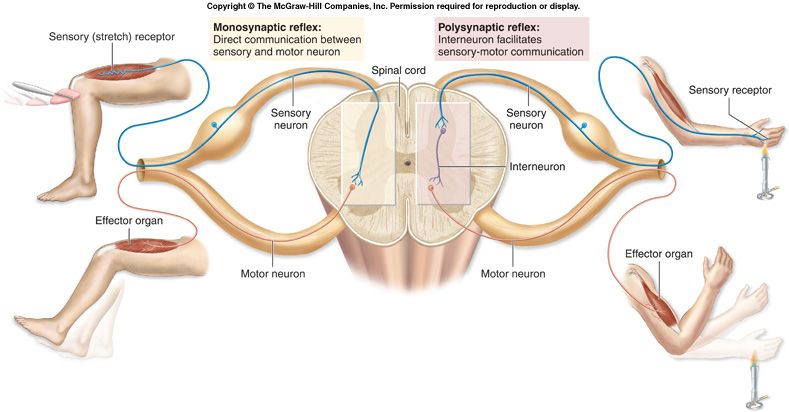
Il riflesso nervoso è una reazione del sistema nervoso centrale a stimolazioni esterne o interne (rilevate tramite i recettori sensoriali), atta a modificare lo stato di contrazione muscolare o di secrezione ghiandolare (reazione motoria, vasomotoria o ghiandolare). I riflessi interessano sia il sistema nervoso volontario o somatico che quello vegetativo (o autonomo). Il meccanismo di base, elementare, di un riflesso nervoso, definito arco riflesso o arco diastaltico, è costituito da: recettore sensoriale > via nervosa afferente o sensitiva > centro nervoso (posto nel sistema nervoso centrale) > via nervosa efferente o motoria > effettore (muscolo o ghiandola). Quando i riflessi nervosi hanno come centro nervoso il midollo spinale, si definiscono riflessi spinali, se i centri sono situati nel tronco encefalico si denominano riflessi troncoencefalici.
I riflessi nervosi vengono suddivisi in semplici (detti anche incondizionati o congeniti) e complessi (o condizionati o acquisiti). Si individuano inoltre due grandi famiglie di riflessi: i riflessi propriocettivi, che originano dalla stimolazione dei propriocettori e che sono monosinaptici (riflesso di stiramento muscolare o riflesso miotatico o osteotendineo ROT, relativo ai fusi neuromuscolari) o disinaptici (riflesso miotatico inverso, relativo agli organi muscolo-tendinei del Golgi), a livello di un solo segmento midollare nel caso dei riflessi spinali, e i riflessi esterocettivi, che derivano dalla stimolazione degli esterocettori della cute e delle mucose, polisinaptici ossia, nel caso dei riflessi spinali, agenti a vari livelli midollari tramite i fasci fondamentali (riflesso plantare, corneale, faringeo, addominale, cremasterico, di retrazione ecc.).
Grazie ai riflessi, in particolare quelli semplici che presentano il grande vantaggio della rapidità (40 m/s per quelli rapidi), i livelli superiori possono demandare a midollo spinale e tronco encefalico, funzioni motorie specifiche che richiedono rapidità di risposta. Tuttavia, grazie alla modalità parallela, i centri encefalici superiori possono interagire direttamente sugli inferiori integrando e vicariando, in maniera immediata, funzioni; questo aspetto risulta fondamentale nel recupero funzionale di alcune lesioni del sistema nervoso centrale; inoltre, ad esempio, il midollo spinale da solo non è in grado di garantire una deambulazione fluida e sicura. I riflessi spinali lavorano in modo gerarchico: controllo dei singoli muscoli, controllo della coordinazione muscolare riguardante una sola articolazione e riguardante più articolazioni.
Fatta eccezione per i riflessi monosinaptici (che derivano sempre dai fusi neuromuscolari), in cui è la stessa la fibra nervosa afferente a connettersi con i motoneuroni alfa, il riflesso spinale avviene in genere grazie all'attività intermediatrice di specifici neuroni, collocati nel midollo spinale, detti interneuroni. Gli interneuroni si inseriscono, tramite sinapsi (connessione fra due neuroni), tra neuroni sensoriali (afferenti dai estero-propriocettori) e motori (efferenti). Gli interneuroni hanno perlopiù azione inibitoria (e talvolta facilitatoria) e sono di tre tipi (classificazione delle fibre nervose): (Ia) che coordinano l'attività di muscoli antagonisti (che esercitano azioni opposte), (Ib) correlati alle afferenze degli organi del Golgi che registrano la tensione tendinea e quindi la contrazione muscolare (da notare che l'influenza di questo tipo di interneurone dipende molto dalle afferenze cutanee a bassa soglia e articolari), cellule di Renshaw che hanno azione inibitoria diretta sul motoneurone. Durante gli atti motori, ogni muscolo che agisce su un'articolazione possiede uno o più muscoli antagonisti che vengono contemporaneamente inibiti (inibizione antagonista) o attivati (attivazione antagonista) tramite rispettivamente un interneurone inibitorio o facilitatorio interposto tra le fibre nervose afferenti (sensoriali) e i relativi motoneuroni alfa.
I riflessi svolgono un ruolo determinante in tutta la fisiologia dell'organismo e naturalmente anche nel sistema dell'equilibrio.
I riflessi spinali semplici vengono infine utilizzati in campo clinico per diagnosticare problematiche neurologiche.
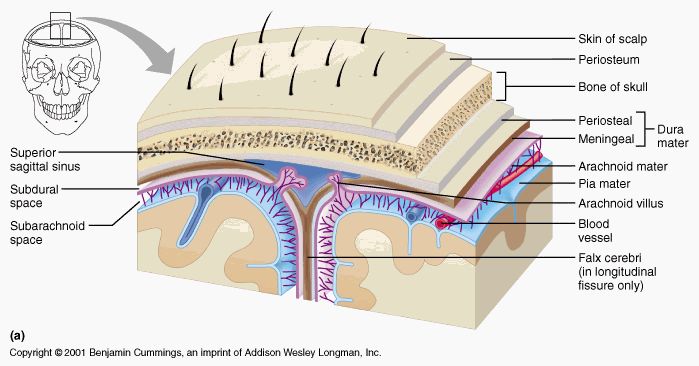 Il sistema nervoso centrale è molto sensibile alla pressione e ancor più alla trazione. Scatola cranica e colonna vertebrale non rappresentano da soli una protezione sufficiente. A tale ulteriore protezione provvedono, all'interno del guscio osseo, uno strato adiposo (presente nello spazio epidurale), le meningi e il liquor cerebrospinale. La dura madre protegge inoltre il sistema nervoso centrale dal diffondersi di infezioni (un'infiammazione intradurale può estendersi all'intero sistema nervoso centrale nel giro di ore o pochi giorni). Le meningi fanno parte del sistema connettivale fasciale costituendo la fascia meningea.
Il sistema nervoso centrale è molto sensibile alla pressione e ancor più alla trazione. Scatola cranica e colonna vertebrale non rappresentano da soli una protezione sufficiente. A tale ulteriore protezione provvedono, all'interno del guscio osseo, uno strato adiposo (presente nello spazio epidurale), le meningi e il liquor cerebrospinale. La dura madre protegge inoltre il sistema nervoso centrale dal diffondersi di infezioni (un'infiammazione intradurale può estendersi all'intero sistema nervoso centrale nel giro di ore o pochi giorni). Le meningi fanno parte del sistema connettivale fasciale costituendo la fascia meningea.
La dura madre e le altre due lamine ad essa concentricamente internamente, aracnoide e pia madre, costituiscono le membrane meningee che rivestono l'intero sistema nervoso centrale. Dura madre e sacco durale inoltre
La dura madre, definita anche pachimeninge per via della sua natura fibrosa e resistente, è la membrana meningea più esterna composta da tessuto connettivale fibroso ricco di fibre elastiche e poco vascolarizzato ed è internamente rivestita da endotelio.
La dura madre intracranica o cefalica aderisce intimamente al periostio del tavolato interno della scatola cranica nella cui cavità si prolunga creando quattro pieghe o sepimenti: tentorio cerebellare (divide trasversalmente la loggia cerebellare da quella cerebrale ossia il cervello o telencefalo dal cervelletto), falce cerebrale o grande falce (piega verticale e mediana situata nella fessura interemisferica dei due emisferi cerebrali), falce cerebellare o piccola falce (piega sagittale posta fra i due lobi cerebellari), diaframma della sella turcica (che chiude superiormente la fossa ipofisaria). Insieme al periostio, la dura madre cefalica costituisce la parete dei seni venosi (rigidi canali in cui circola il sangue venoso).
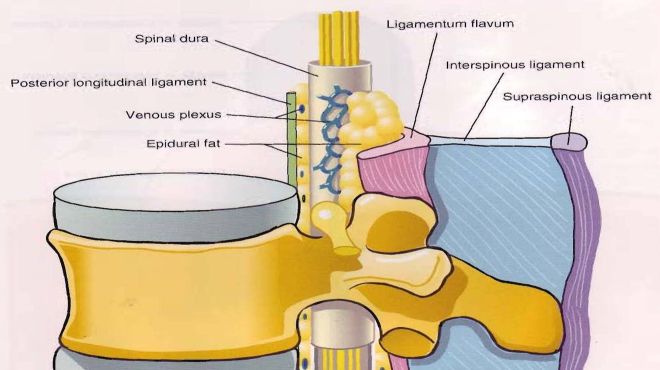 La dura madre cefalica si continua con la dura spinale, a livello del forame magno (forame occipitale), dove i suoi foglietti periosteo e meningeo, fusi a livello del cranio, si sdoppiano costituendo rispettivamente il rivestimento periostale interno delle vertebre (dello speco vertebrale) e il sacco durale separato dalle pareti interne del canale vertebrale dallo spazio epidurale. Questo spazio peridurale risulta riempito da tessuto adiposo (tendenzialmente molle e globulare e la cui quantità è in rapporto a quella di tutto l’organismo, per cui è maggiore nei soggetti obesi), da frange di tessuto connettivo e da vasi sanguigni e linfatici. Lo spazio epidurale è chiuso caudalmente a livello del coccige tramite la membrana sacro-coccigea, è a contatto anteriormente con il legamento longitudinale posteriore, lateralmente con i forami intervertebrali e le lamine vertebrali, posteriormente con i legamenti gialli (flavum). Esso pertanto non è uno spazio chiuso in quanto comunica tramite i forami intervertebrali con i tessuti perivertebrali. Il suo spessore è in rapporto alla posizione e alle dimensioni del sacco durale;:essendo quest'ultimo posto più vicino alla parete anteriore del canale vertebrale, lo spazio peridurale è maggiore sulla linea posteriore mediana specie a partire dalla seconda vertebra lombare (qui misura circa 5-6 mm nell’adulto) in giù, dove il sacco durale comincia a ridursi di diametro. Nella regione medio toracica tale distanza, sempre nell'adulto, è di circa 3-4 mm, nella regione cervicale inferiore è di circa 1,5- 2 mm. All’interno vengono iniettate e si diffondono le soluzione anestetiche.
La dura madre cefalica si continua con la dura spinale, a livello del forame magno (forame occipitale), dove i suoi foglietti periosteo e meningeo, fusi a livello del cranio, si sdoppiano costituendo rispettivamente il rivestimento periostale interno delle vertebre (dello speco vertebrale) e il sacco durale separato dalle pareti interne del canale vertebrale dallo spazio epidurale. Questo spazio peridurale risulta riempito da tessuto adiposo (tendenzialmente molle e globulare e la cui quantità è in rapporto a quella di tutto l’organismo, per cui è maggiore nei soggetti obesi), da frange di tessuto connettivo e da vasi sanguigni e linfatici. Lo spazio epidurale è chiuso caudalmente a livello del coccige tramite la membrana sacro-coccigea, è a contatto anteriormente con il legamento longitudinale posteriore, lateralmente con i forami intervertebrali e le lamine vertebrali, posteriormente con i legamenti gialli (flavum). Esso pertanto non è uno spazio chiuso in quanto comunica tramite i forami intervertebrali con i tessuti perivertebrali. Il suo spessore è in rapporto alla posizione e alle dimensioni del sacco durale;:essendo quest'ultimo posto più vicino alla parete anteriore del canale vertebrale, lo spazio peridurale è maggiore sulla linea posteriore mediana specie a partire dalla seconda vertebra lombare (qui misura circa 5-6 mm nell’adulto) in giù, dove il sacco durale comincia a ridursi di diametro. Nella regione medio toracica tale distanza, sempre nell'adulto, è di circa 3-4 mm, nella regione cervicale inferiore è di circa 1,5- 2 mm. All’interno vengono iniettate e si diffondono le soluzione anestetiche.
Il sacco durale rappresenta un lungo manicotto che circonda il midollo spinale e il midollo sacrale, possiede delle estensioni tubulari laterali che rivestono le radici dei nervi spinali, accompagnandoli attraverso i fori di coniugazione intervertebrali. Nel punto in cui la radice nervosa dorsale e quella ventrale si uniscono per formare il nervo spinale, la dura madre si fonde in un singolo manicotto (includente anche aracnoide e pia madre) che si continua come epinevrio del nervo spinale. Caudalmente, il sacco durale si restringe, a livello della II-III vertebra sacrale, riveste la cauda equina e il midollo sacrale (cisterna lombare) nonchè il filum terminale ancorando quest'ultimo al periostio coccigeo (faccia dorsale della I-II vertebra coccigea) tramite il legamento coccigeo formato dalle tre membrane meningee fuse assieme. Il sacco durale contiene quindi aracnoide, pia madre, midollo spinalemidollo spinale, midollo sacrale, radici nervose spinali, nervi spinali e della cauda equina e liquor cerebrospinale.
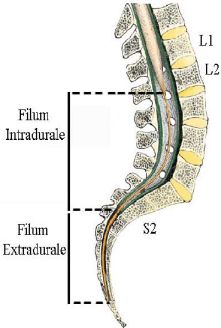 Il filum o filo terminale spinale rappresenta il prolungamento terminale del midollo spinale (contenente il midollo sacrale) costituito da una sottile e lunga (diametro ca. 2 mm, lunghezza totale ca. 25 cm) struttura fibrovascolare, composta principalmente da tessuto fibroso in continuità con quello della pia madre. Il filum terminale ancora il cono midollare, al periostio del coccige.
Il filum o filo terminale spinale rappresenta il prolungamento terminale del midollo spinale (contenente il midollo sacrale) costituito da una sottile e lunga (diametro ca. 2 mm, lunghezza totale ca. 25 cm) struttura fibrovascolare, composta principalmente da tessuto fibroso in continuità con quello della pia madre. Il filum terminale ancora il cono midollare, al periostio del coccige.
E' possibile suddividere strutturalmente Il filum terminale in due porzioni:
1) filo terminale interno, contenuto entro la guaina tubulare del sacco durale, circondato dai nervi della cauda equina (grazie al proprio colore bluastro-bianco il filum terminale può essere facilmente distinto dalla cauda equina), si estende per ca. 15 cm dal cono midollare al bordo inferiore della seconda vertebra sacrale (S2), dove si fonde con il sacco durale stesso.
2) filo terminale esterno, dalla punta del sacco durale prosegue caudalmente fino al livello della I o II vertebra coccigea rivestito dalle tre membrane meningee fuse tra loro che lo ancorano al periostio della relativa faccia dorsale formando il legamento coccigeo (o sacro coccigeo).
L’interesse attorno al filum terminale è perlopiù dovuto alla "sindrome del midollo spinale ancorato" (tethered cord syndrome), in cui il midollo spinale è trazionato inferiormente da un filum patologico. La presenza infatti di una quantità elevata di fibre connettivali elastiche disposte longitudinalmente all’interno del filum terminale gli consentono un ruolo di "ammortizzatore" fisiologico atto a prevenire eccessive trazioni midollari durante i normali movimenti del tronco (flessione antero-posteriore e rotazione).
Va inoltre considerato che a contatto con la superficie esterna del filo terminale sono presenti alcune fibre nervose, probabilmente nervi coccigei rudimentali. Infine, il canale centrale del midollo spinale prosegue al suo interno per 5 o 6 centimetri.
Oltre che dal filum terminale, il midollo spinale è mantenuto in posizione fisiologica dai legamenti dentati che lo ancorano bilateralmente al sacco durale.
Situata direttamente all'interno della dura madre cefalica e del sacco durale (ai quali è mantenuta adesa tramite la pressione del liquor cerebrospinale, presente nello spazio subaracnoideo, e dai legamenti dentati che si dipartono dalla pia madre), l’aracnoide è una sottile e trasparente membrana, non vascolarizzata, composta da fibre collagene incrociate a mo di fitta ragnatela (da cui il nome). La membrana aracnoidea, a livello dei seni venosi cerebrali, presenta le granulazioni o granuli di Pacchioni o villi aracnoidei, formati da tessuto trabecolare subaracnoideo rivestito dall'aracnoide (assenti nei bambini, si sviluppano verso il 10°-12° anno aumentando in special modo in età senile, allorché spesso si calcificano) che riassorbono nel circolo venoso il liquido cerebrospinale. L'aracnoide è saldamente connessa alla membrana più interna, la pia madre, tramite tralci connettivali (aracnoide e pia madre vengono definite leptomeninge, data la loro sottigliezza).
La cavità subaracnoidea, definita spazio subaracnoideo o subaracnoidale, spessa alcuni mm che si forma tra la membrana aracnoidea e la pia madre, è ripiena di liquido cefalorachidiano (liquor cerebrospinale). Lo spazio subaracnoideo presenta delle porzioni espanse definite cisterne liquorali o della base (essendo perlopiù presenti nella base del cranio). Tali cisterne si determinano in quanto mentre la pia madre riveste intimamente l'encefalo, seguendo tutte le sue scissure e solchi, l'aracnoide e la dura madre vi possono passare a ponte, formando così le dilatazioni dello spazio subaracnoideo.
Le cavità cerebrali e midollari contenenti liquor cerebrospinale servono a proteggere da sollecitazioni meccaniche e a consentire espansioni limitate dell'encefalo (ad es. dopo una commozione cerebrale) senza aumento della pressione endocranica.
La pia madre, costituita da fibre connettive reticolari ed elastiche, rappresenta la membrana meningea più interna, più sottile e più vascolarizzata (la maggior parte dei suoi vasi irrorano le strutture nervose sottostanti). Essa aderisce, penetrando in tutte le infrattuosità, alla superficie dell’encefalo (molto intimamente), del midollo spinale e sacrale. Dalla superficie della pia madre e per tutta la sua lunghezza si dipartono, a destra e sinistra sul piano frontale, i legamenti dentati, lamine festonate che fissano il midollo spinale alla parete del sacco durale, sospendendolo in modo elastico, tramite prolungamenti a punta (18-22 per lato con l'apice inguainato dall'aracnoide) sottendenti delle arcate al di sotto delle quali le radici ventrali e dorsali dei nervi spinali si incontrano.
Insieme alla membrana aracnoidea e al sacco durale, la pia madre riveste il filum terminale (formando anche il legamento coccigeo che lo ancora al coccige) e le radici dei nervi spinali fino a costituire parte dei rivestimenti epineurali.
La pia madre forma i plessi corioidei dei ventricoli cerebrali, granulazioni rosee riccamente irrorate e ricoperte da epitelio ependimale (costituente la barriera emato-liquorale), che producono il liquor cefalorachidiano.
Il liquor cerebro-spinale (liquido cefalo-rachidiano) è un ultrafiltrato incolore e trasparente del plasma sanguineo, con esso isotonico ma di differente composizione (povero di linfociti e proteine, con molte meno albumine, glucosio, potassio, calcio), che permea il sistema nervoso centrale. Il liquor cerebrospinale si trova nello spazio subaracnoideo (tra aracnoide e pia madre), bagna l'encefalo, il midollo spinale, le radici e i relativi nervi periferici e i globi oculari. Il liquido cefalorachidiano occupa anche gli spazi "interni" del sistema nervoso centrale quali le cisterne, i ventricoli cerebrali e il sottile (e spesso occluso) canale midollare centrale (spazio liquorale interno).
Il liquor cefalorachidiano ha funzione di protezione e sostegno (l'encefalo ci galleggia letteralmente diminuendo il suo peso a 25 grammi). Esso è in grado di assorbire traumi esterni distribuendone le relative forze creando, in condizioni fisiologiche, l’ambiente ottimale per le attività delle cellule nervose: chimicamente la sua composizione è simile a quella del liquido interstiziale del tessuto nervoso, vi è quindi uno scambio continuo tra essi, parte dei metaboliti anzichè essere riassorbiti dai capillari finisce nei ventricoli e poi nel liquor (pertanto ogni alterazione nell'interstizio si trasmette nel liquor, da ciò deriva la diagnostica liquorale), supplisce alla quasi totale assenza di un sistema linfatico del sistema nervoso centrale, trasporta messaggi ormonali. Inoltre, il liquido cefalo-rachidiano spinge l’aracnoide perifericamente mantenendola adesa alla soprastante dura madre.
La trasmissione degli impulsi nel sistema nervoso avviene quindi tramite specifici messaggeri che viaggiano lungo le vie neurali così come per mezzo del sangue, della linfa, della matrice extracellulare e del liquor cerebrospinale.
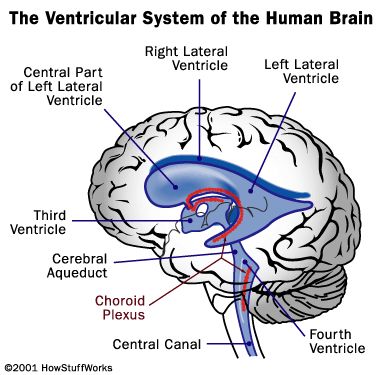 La produzione di liquido cefalorachidiano è di tipo attivo (è indipendente dalla pressione arteriosa) ed è pari a circa 300-500 ml al giorno con un ricambio giornaliero di tre-quattro volte; pertanto il volume di liquor circolante si aggira sui 100-200 ml. Il liquido cerebrospinale viene prodotto dai plessi corioidei situati nei ventricoli cerebrali rappresentati da quattro cavità comunicanti presenti all'interno dell'encefalo (residuo del lume del primitivo tubo neurale a livello encefalico) in cui viene prodotto e circola il liquido cefalorachidiano. Il primo e il secondo, definiti ventricoli laterali, si trovano all'interno degli emisferi cerebrali (di gran lunga quelli di maggiori dimensione, si estendono dal lobo frontale ai lobi occipitali) e comunicano, tramite i fori interventricolari destro e sinistro di Monro, con il terzo ventricolo, impari e mediano, contenuto nel diencefalo il quale, a sua volta, è in comunicazione per mezzo dell'acquedotto mesencefalico del Silvio (o acquedotto cerebrale) col quarto ventricolo presente nel romboencefalo. Il liquido cerebrospinale si riversa dal quarto ventricolo nello spazio subaracnoideo attraverso il foro di Magendie e i due fori di Luschka, a livello del midollo allungato.
La produzione di liquido cefalorachidiano è di tipo attivo (è indipendente dalla pressione arteriosa) ed è pari a circa 300-500 ml al giorno con un ricambio giornaliero di tre-quattro volte; pertanto il volume di liquor circolante si aggira sui 100-200 ml. Il liquido cerebrospinale viene prodotto dai plessi corioidei situati nei ventricoli cerebrali rappresentati da quattro cavità comunicanti presenti all'interno dell'encefalo (residuo del lume del primitivo tubo neurale a livello encefalico) in cui viene prodotto e circola il liquido cefalorachidiano. Il primo e il secondo, definiti ventricoli laterali, si trovano all'interno degli emisferi cerebrali (di gran lunga quelli di maggiori dimensione, si estendono dal lobo frontale ai lobi occipitali) e comunicano, tramite i fori interventricolari destro e sinistro di Monro, con il terzo ventricolo, impari e mediano, contenuto nel diencefalo il quale, a sua volta, è in comunicazione per mezzo dell'acquedotto mesencefalico del Silvio (o acquedotto cerebrale) col quarto ventricolo presente nel romboencefalo. Il liquido cerebrospinale si riversa dal quarto ventricolo nello spazio subaracnoideo attraverso il foro di Magendie e i due fori di Luschka, a livello del midollo allungato.
Il liquor cerebrospinale possiede movimenti dinamici propri ma ritmati dall'attività cardiaca. Durante la contrazione sistolica il liquor dai ventricoli si dirige verso gli spazi oculari, intrarachidei e nel canale midollare centrale (quest'ultimo si espande in alto nei ventricoli cerebrali); durante il rilassamento diastolico il senso di flusso si inverte risalendo senza però oltrepassare i forami di Magendie e Luschka in cui la direzione è sempre dal comparto intracranico a quello extracranico.
Una volta svolto il suo compito il liquor viene riassorbito nel circolo venoso perlopiù a livello delle granulazioni o villi aracnoidei (granulazioni di Pacchioni) che sporgono all'interno dei seni venosi cerebrali (in particolar modo nel seno sagittale). Granulazioni sono presenti anche vicino alla radice nervosa dorsale del gangli spinali lungo tutta la colonna. Un ulteriore riassorbimento avviene a livello della porzione meningea dell'epinevrio dei nervi spinali (questo spiega come un liquido introdotto nello spazio subaracnoideo si diffonda in tutta la guaina del nervo).
Se il deflusso del liquido cefalorachidiano è ostacolato, si registra un pericoloso aumento della pressione encefalica che può comportare conseguenze anche molto gravi (fino al decesso). L'idrocefalo è quella condizione per la quale il liquor cerebrospinale si accumula nelle cavità ventricolari, che si dilatano e determinano un aumento della pressione endocranica.
L'analisi cellulare, biochimica e microbiologica del liquor è in grado di dimostrare o escludere soprattutto processi infiammatori o infettivi del sistema nervoso.
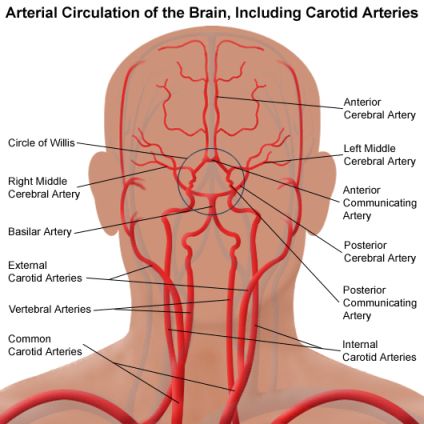
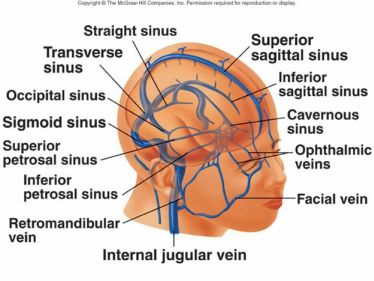 Relativamente alla vascolarizzazione del sistema nervoso centrale, l'elevato fabbisogno energetico fa sì che l'encefalo sia molto sensibile alle variazioni di irrorazione sanguinea. La situazione è particolarmente delicata in quanto il collo, attraverso il quale corrono le arterie che si distribuiscono nell'encefalo, è particolarmente mobile e sottoposto, col passare degli anni, a stress e conseguenti alterazioni muscolo-scheletriche di orgine posturale e/o traumatica; può quindi succedere che una grossa arteria possa subire compressioni con danni consegurenti a tutto il sistema nervoso.
Relativamente alla vascolarizzazione del sistema nervoso centrale, l'elevato fabbisogno energetico fa sì che l'encefalo sia molto sensibile alle variazioni di irrorazione sanguinea. La situazione è particolarmente delicata in quanto il collo, attraverso il quale corrono le arterie che si distribuiscono nell'encefalo, è particolarmente mobile e sottoposto, col passare degli anni, a stress e conseguenti alterazioni muscolo-scheletriche di orgine posturale e/o traumatica; può quindi succedere che una grossa arteria possa subire compressioni con danni consegurenti a tutto il sistema nervoso.
L'encefalo e le meningi cerebrali vengono rifornite essenzialmente dalle due arterie carotidi interne e dell'arteria basilare che deriva dalla confluenza, anteriormente al ponte, delle due arterie vertebrali che attraversano le apofisi trasverse delle vertebre cervicali. L'arteria carotide esterna fornisce rami meningei nella dura madre.
Fra la superficie interna della scatola cranica e la dura madre si ritrovano i seni venosi, rigidi canali a sezione triangolare dove si riversa il sangue refluo. Tra i numerosi seni venosi, i sagittali (superiore e inferiore), situati centralmente, si dividono simmetricamente nei due seni sigmoidei. Il seno cavernoso, posto lateralmente alla sella turcica (presente alla base del cranio e contenente l'ipofisi), riceve dalla vena oftalmica il sangue proveniente dalle regioni anteriore e inferiore dell'encefalo e da alcune regioni del volto (processi suppurativi della cute del viso possono comportare pericolose infezioni dei seni venosi). Le due vene giugulari interne rappresentano le normali vie di deflusso e veicolano il sangue venoso intracranico nelle vene succlavie (posteriormente all'articolazione sterno-clavicolare) e quindi nella vena cava superiore.
In posizione seduta o eretta, i seni venosi presentano una pressione negativa trovandosi ca. 30 cm più in alto dell'atrio destro del cuore. Essi quindi, analogamente alle grosse vene del collo, devono necessariamente mantenersi pervi per non collassare.
A causa della rigida struttura dei seni venosi della dura madre, un violento trauma, capace di spostare bruscamente il contenuto craniale contro di essa, può comportare la rottura per pressione delle vene cerebrali creando una pericolosa emorragia che spesso si sviluppa dopo alcuni giorni.
La circolazione linfatica della dura madre avviene attraverso piccoli canali comunicanti che raggiungono gli spazi epidurali e sottodurali.
Il midollo spinale presenta una ricca vascolarizzazione alimentata dalle arterie radicolari che prendono origine: nel collo dalle arterie cervicale ascendente e vertebrale, nel torace dalle arterie intercostali, nella zona lombare dalle arterie lombari, nella regione sacrale da quelle sacrali. Le arterie radicolari attraversano i fori di coniugazione vertebrali e formano, in corrispondenza della porzione superiore del midollo spinale, due tronchi arteriosi posteriori e uno anteriore (arterie spinali posteriori e arteria spinale anteriore) che si ramificano all'interno della pia madre penetrando nel midollo spinale a raggiera.
Il sistema venoso del midollo spinale è molto simile a quello arterioso (le vene sono perlopiù satelliti delle arterie): dalla sostanza grigia e dalla bianca origina le vene intramidollari che radialmente convogliano in un'ampia rete perimidollare che si raccoglie in sei collettori longitudinali da cui nascono le vene radicolari che raggiungono i fori intervertebrali.
Non sono presenti vasi linfatici.
L'endotelio dei vasi capillari del sistema nervoso centrale si presenta continuo (non fenestrato) con cellule unite da giunzioni occludenti o serrate (tight junction). Ciò crea una barriera altamente selettiva, definita barriera emato-encefalica, che impedisce il transito passivo di sostanze idrofile e/o con grande peso molecolare dal flusso sanguigno all'interstizio e quindi ai neuroni. Ulteriore fattore che contribuisce alla formazione della barriera emato-encefalica è la presenza di cellule della glia (tra cui gli astrociti) che con i propri tipici prolungamenti a raggiera formano una guaina completa intorno ai capillari fungendo da ostacolo secondario al passaggio di molecole idrofile.
In condizioni fisiologiche tale barriera possiede la capacità di proteggere il sistema nervoso centrale da elementi nocivi (sostanze estranee, cellule, batteri, virus, farmaci, ormoni e neurotrasmettitori diretti ad altri organi ecc.) presenti nel sangue pur tuttavia permettendo il passaggio di sostanze necessarie alle funzioni metaboliche così da mantenere l'omeostasi del sistema nervoso centrale.
Sostanze lipofile come l’ossigeno, il diossido di carbonio, l’etanolo e gli ormoni steroidi sono in grado di attraversare passivamente la membrana emato-encefalica mentre altre quali aminoacidi, glucosio, insulina, lipoproteine a bassa densità (LDL) ecc. richiedono un trasporto attivo mediante specifiche vescicole.
L'efficacia della barriera emato-encefalica può essere compromessa in situazioni quali ipertensione arteriosa, iperosmolarità, infezioni, stati infiammatori, traumi, esposizioni a radiazioni ecc.).
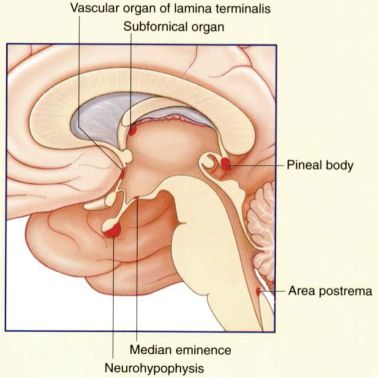 La barriera emato-encefalica non è presente a livello di alcune aree dell'encefalo poste nei pressi del sistema ventricolare, definite "organi circumventricolari", in cui, grazie al facile transito di molecole dal sangue ai ventricoli e viceversa, il sistema nervoso centrale monitora la composizione del sangue circolante. Organi circumventricolari" sono: la neuroipofisi, l’eminenza mediana dell’ipofisi, l’epifisi, l'area postrema (“centro del vomito”), l'organo subfornicale (regola i liquidi corporei), l'organo vascoloso della lamina terminale,
La barriera emato-encefalica non è presente a livello di alcune aree dell'encefalo poste nei pressi del sistema ventricolare, definite "organi circumventricolari", in cui, grazie al facile transito di molecole dal sangue ai ventricoli e viceversa, il sistema nervoso centrale monitora la composizione del sangue circolante. Organi circumventricolari" sono: la neuroipofisi, l’eminenza mediana dell’ipofisi, l’epifisi, l'area postrema (“centro del vomito”), l'organo subfornicale (regola i liquidi corporei), l'organo vascoloso della lamina terminale,
Come gli organi circumventricolari, anche i plessi corioidei presentano un normale endotelio fenestrato (senza giunzioni serrate), al loro esterno esiste però un sottile epitelio (epitelio ependimale o ependima) formato da specifiche cellule della glia definite ependimali (o ependimociti). L'ependima delimita le cavità del sistema nervoso centrale, rivestendo i ventricoli cerebrali e il canale centrale del midollo spinale, e costituisce una seconda barriera (oltre a quella emato-encefalica), la barriera emato-liquorale. Questa barriera emato-fluido cerebrospinale seleziona (in base alla liposolibilità, grandezza e tramite trasporto attivo per le sostanze idrosolubili) il passaggio di sostanze dal plasma al liquor cerebrospinale e viceversa attraverso meccanismi di riassorbimento, supplendo in tal modo alla carenza di vasi linfatici del sistema nervoso centrale. Inoltre, il battito delle ciglia delle cellule ependimali favorisce la circolazione del liquido cerebrospinale.
Essendo le retine oculari estensioni del sistema nervoso centrale, va qui considerata anche la barriera emato-retinica, facente parte della barriera emato-oculare e consistente in un endotelio monostrato continuo e con giunzioni occludenti (tight junction) con all'esterno un epitelio pigmentato (barriera emato-oculare esterna). La barriera emato-retinica regola il passaggio di molecole dai vasi retinici (coriocapillari) alla retina (la retinopatia diabetica consiste nell'indebolimento fino al collasso della barriera emato-retinica).
Pertanto le barriere emato-encefalica, emato-liquorale ed emato-retinica assicurano l'omeostasi dei liquidi extracellulari nei quali operano e vivono i neuroni.
- Stress e Benessere, Giovanni Chetta
- Alimentazione e Benessere, Giovanni Chetta
- Postura e Benessere, Giovanni Chetta
- Il Sistema Connettivo, Giovanni Chetta
- Dalla MEC alla postura, Giovanni Chetta
- Ginnastica Posturale TIB, Giovanni Chetta
- Il Massaggio Antistress TIB, Giovanni Chetta
- Massaggio & Bodywork TIB, Giovanni Chetta
