Approfondimenti |
|

La propriocezione rappresenta la capacità del sistema nervoso centrale di percepire la posizione del corpo e delle sue parti nonchè della contrazione muscolare e del movimento nello spazio dei diversi distretti corporei (cinestesia) anche senza l'apporto della vista. Tale sensibilità insorge all'interno di un organo (fascia connettivale, articolazioni, tendini, muscoli, capsule articolari, cute, visceri ecc.), attraverso la stimolazione di specifici recettori sensoriali: esterocettori cutanei, propriocettori, enterocettori e recettori vestibolari.
La propriocezione assume quindi un'importanza fondamentale nel complesso sistema dell'equilibrio e del movimento (l'atassia rappresenta clinicamente l'alterazione della propriocezione determinante la progressiva perdita della coordinazione muscolare e quindi motoria).
Dai recettori periferici (inclusi gli esterocettori retinici degli occhi) dipende la conoscenza su quale è la nostra "conformazione" e posizione spaziale; in qualche misura, per rispondere alla domanda "chi sono io?", occorre anche rispondere alla domanda "dove sono io?".
E' dai riflessi estero-propriocettivi che nascono le rappresentazioni mentali (engrammi) che consentono la nascita di abilità motorie complesse ma non solo, azioni e movimenti hanno un ruolo centrale nei processi di rappresentazione mentale, a partire dalla fase embrionale. L'embrione, infatti, è innanzitutto un organismo motorio. Nella fase embrionale, in quella fetale e in quella della prima infanzia, l'azione precede la sensazione: vengono compiuti dei movimenti riflessi e poi se ne ha la percezione a livello cerebrale. Le funzioni motorie e il corpo, spesso considerati entità inferiori e subordinate alle attività cognitive sono invece all'origine di esse, incluso lo stesso linguaggio che forma la nostra mente e nostri pensieri. La propriocezione rappresenta quindi "la coscienza di sè", perdere il controllo sul proprio corpo significa, di conseguenza, perdere il controllo sui propri pensieri ed emozioni. "Essere" e "funzionare" sono inscindibili.
"Muscoli, tendini, capsule, legamenti sono organi di senso e organi di moto, impegnati nella ricezione della stimolazione ambientale prima che nell'effettuazione della risposta di moto...il senso non può giammai dissociarsi dal moto" (R. Paparella Teccia, 1988).
Normalmente la quantità di fibre nervose sensitive presenti in un nervo supera di gran lunga quelle motorie (nel nervo tibiale ad es. la proporzione fra esse è di ca. 3 a 1), ciò sta anche a indicare l'importanza che l'organismo riserva al sistema di feed-back. Inoltre ricerche sulla corteccia motoria hanno dimostrato che essa è organizzata non tanto in base alle aree topografiche corporee quanto piuttosto relativamente a specifici movimenti corporei complessi indirizzati nello spazio verso un obiettivo definito. Da ciò ne consegue che un movimento eseguito immaginando (visualizzando), ad es., di afferrare, respingere o disegnare un oggetto coinvolge il sistema nervoso molto più dello stesso gesto eseguito solo meccanicamente, stimolando e sviluppando in tal modo la propriocezione di quella specifica articolazione o area corporea; ciò risulta ovviamente importante ai fini rieducativi.
I recettori sensoriali nervosi sono formazioni distribuite in tutto il corpo con la funzione di ricevere i diversi stimoli esterni e interni all'organismo e di trasmetterli, trasducendoli in impulsi elettrici, attraverso le fibre nervose afferenti, al sistema nervoso centrale. I recettori nervosi vengono classificati in vari modi. Alcuni di essi formano strutture complesse tali da considerarsi organi propriamente detti di senso; ad es. l'occhio e l'orecchio.
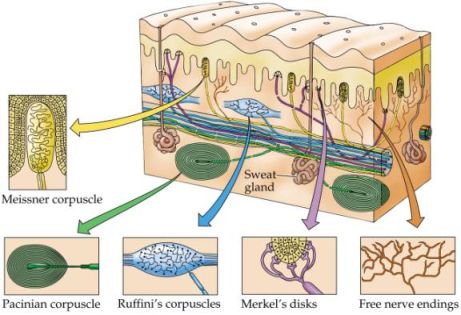 Fra i neuroreccettori sensoriali, gli esterocettori cutanei e i propriocettori importanti dal punto di vista posturologico sono recettori sensoriali sensibili a stimoli provenienti dall'esterno quali il tatto, la pressione cutanea e altre variazione strutturali di tipo meccanico (meccanocettori) di derivazione anche interna. Essi possiedono caratteristiche strutturali e di adattamento diverse e suscitano variazioni organiche locali e in taluni casi anche generali. Tali recettori sensoriali sono responsabili della propriocezione e cinestesia fornendo al sistema dell'equilibrio informazioni sull'ambiente (esterno e interno) consentendo al sistema posturale di conoscere, istante per istante, la posizione e lo stato di ogni osso, muscolo, organo e sistema.
Fra i neuroreccettori sensoriali, gli esterocettori cutanei e i propriocettori importanti dal punto di vista posturologico sono recettori sensoriali sensibili a stimoli provenienti dall'esterno quali il tatto, la pressione cutanea e altre variazione strutturali di tipo meccanico (meccanocettori) di derivazione anche interna. Essi possiedono caratteristiche strutturali e di adattamento diverse e suscitano variazioni organiche locali e in taluni casi anche generali. Tali recettori sensoriali sono responsabili della propriocezione e cinestesia fornendo al sistema dell'equilibrio informazioni sull'ambiente (esterno e interno) consentendo al sistema posturale di conoscere, istante per istante, la posizione e lo stato di ogni osso, muscolo, organo e sistema.
- Recettori interstiziali, perlopiù terminazioni nervose libere (non capsulate) sparse ovunque nella cute, nei tessuti connettivi e negli interstizi muscolari (esterecettori cutanei e propriocettori). Questi piccoli recettori, praticamente presenti ovunque nell'organismo (meningi e legamenti viscerali inclusi e massima concentrazione nel periostio) rappresentano la famiglia di recettori nervosi di gran lunga più numerosa. Ciò che occorre considerare è che nell’innervazione muscolare dell'uomo le fibre sensitive derivano solo per ca. il 25% dai ben noti recettori del Golgi, Ruffini, Pacini e Paciniformi (fibre tipo I e II) mentre tutta la restante parte ha origine dai “recettori interstiziali” (fibre tipo III e IV). Circa il 90% dei recettori interstiziali sono demienilizzati (tipo IV) mentre il restanti posseggono una sottile guaina mielinica (tipo III). Ad azione più lenta rispetto ai recettori tipo I e II, in passato sono stati considerati perlopiù nocicettori, termo e chemiorecettori (sensibili a vibrazioni con frequenze inferiori ai 100 Hz). In realtà molti di loro risultano multimodali e in maggioranza meccanorecettori (sensibili a tatto, pressione e trazione) suddivisibili in due sottogruppi, di uguale numero circa, in base alla loro soglia di attivazione tramite stimoli pressori: low-treshold (LTP) e high-treshold pressure (HTP). Sono presenti recettori interstiziali a lento o a rapido adattamento. L'attivazione, in determinati stati patologici di recettori interstiziali sensibili sia a stimoli dolorifici che meccanici (in maggioranza HTP) può generare sindromi dolorose in assenza delle classiche irritazioni nervose (es. compressioni radicolari). Questo network sensoriale, oltre ad avere una funzione di rilevamento afferente del posizionamento e del movimento dei segmenti corporei, influenza, per mezzo di intime connessioni, il sistema nervoso autonomo riguardo funzioni quali la regolazione della pressione sanguinea, del battito cardiaco e della respirazione sintonizzandole, in maniera molto precisa, alle esigenze tissutali locali. L'attivazione dei meccanorecettori interstiziali agisce sul sistema nervoso autonomo inducendolo a variare la pressione locale di arteriole e capillari presenti nel tessuto connettivo, influenzando così il passaggio di plasma dai vasi alla matrice extracellulare variandone quindi la viscosità locale. Inoltre la stimolazione tramite pressioni lente e profonde dei recettori interstiziali, così come quella dei recettori di Ruffini, è in grado di incrementare il tono vagale (sistema nervoso parasimpatico) generando cambiamenti organici globali comportanti un profondo e benefico rilassamento.
- Recettori dei follicoli piliferi (o organi terminali piliferi), esterocettori cutanei costituiti da fibre nervose sensitive non capsulate avvolte a spirale attorno alla radice del pelo cutaneo, al cui movimento sono particolarmente sensibili, tramite rapido adattamento.
 - Cellule o dischi di Merkel (recettori SAI), esterocettori cutanei coastituiti da grosse cellule (ricche di mitocondri) a terminazione espansa non capsulata connessa con una fibra nervosa mielinica di grande diametro (tipo A beta - tipo II), a lento adattamento e bassa soglia di attivazione, situati in corrispondenza delle creste epidermiche, al di sotto dello strato basale, spesso raggruppati a costituire il recettore a cupola di Iggo estremamente sensibile. Le cellule di Merkel insieme ai corpuscoli di Meissner svolgono un ruolo importante nel determinare le caratteristiche spaziali delle sensazioni tattili. Essendo le grosse cellule di Merkel sono sensori proporzionali (sensori P) in quanto trasformano una pressione doppia sulla cute in circa un numero doppio di potenziali d'azione al secondo. Poichè trasmettono gli impulsi tramite una singola fibra nervosa, solo l'eccitazione contemporanea di più cellule di Merkel adiacenti viene trasmessa come segnale al midollo spinale (convergenza di segnali).
- Cellule o dischi di Merkel (recettori SAI), esterocettori cutanei coastituiti da grosse cellule (ricche di mitocondri) a terminazione espansa non capsulata connessa con una fibra nervosa mielinica di grande diametro (tipo A beta - tipo II), a lento adattamento e bassa soglia di attivazione, situati in corrispondenza delle creste epidermiche, al di sotto dello strato basale, spesso raggruppati a costituire il recettore a cupola di Iggo estremamente sensibile. Le cellule di Merkel insieme ai corpuscoli di Meissner svolgono un ruolo importante nel determinare le caratteristiche spaziali delle sensazioni tattili. Essendo le grosse cellule di Merkel sono sensori proporzionali (sensori P) in quanto trasformano una pressione doppia sulla cute in circa un numero doppio di potenziali d'azione al secondo. Poichè trasmettono gli impulsi tramite una singola fibra nervosa, solo l'eccitazione contemporanea di più cellule di Merkel adiacenti viene trasmessa come segnale al midollo spinale (convergenza di segnali).
 - Corpuscoli ramificati e lanceolati di Meissner (recettori RA), esterocettori cutanei (situati fra epidermide e derma) tattili di particolare sensibilità, costituiti da una terminazione nervosa capsulata (capsula di tessuto connettivo) e allungata in rapporto con una tortuosa fibra nervosa mielinica di grande diametro (tipo A beta - tipo II), a rapido adattamento. Tali corpuscoli pertanto già dopo poco tempo non reagiscono a una pressione persistente, ma solo a sue variazioni: quanto più rapidamente varia la pressione, tanto più intensamente scarica la fibra nervosa (sensori differenziali o sensori D). I corpuscoli di Meissner sono molto numerosi nelle regione cutanee glabre, ossia senza peli (specie in polpastrelli, labbra, piedi ecc.). Essi infatti vengono, ad es., utilizzati per afferrare con precisione oggetti di piccole dimensioni.
- Corpuscoli ramificati e lanceolati di Meissner (recettori RA), esterocettori cutanei (situati fra epidermide e derma) tattili di particolare sensibilità, costituiti da una terminazione nervosa capsulata (capsula di tessuto connettivo) e allungata in rapporto con una tortuosa fibra nervosa mielinica di grande diametro (tipo A beta - tipo II), a rapido adattamento. Tali corpuscoli pertanto già dopo poco tempo non reagiscono a una pressione persistente, ma solo a sue variazioni: quanto più rapidamente varia la pressione, tanto più intensamente scarica la fibra nervosa (sensori differenziali o sensori D). I corpuscoli di Meissner sono molto numerosi nelle regione cutanee glabre, ossia senza peli (specie in polpastrelli, labbra, piedi ecc.). Essi infatti vengono, ad es., utilizzati per afferrare con precisione oggetti di piccole dimensioni.
- Corpuscoli ramificati di Ruffini (recettori SAII), costituiti da terminazioni nervose capsulate e pluriramificate, dove termina una fibra nervosa mielinica di grande diametro (tipo A beta - tipo II) e numerose fibre connettivali, presenti negli strati cutanei e tissutali profondi, nelle capsule articolari (strati superficiali), legamenti, dura madre ecc.. I corpuscoli di Ruffini sono pertanto sia estrocettori cutanei che propriocettori. Essi reagiscono maggiormente agli stimoli veloci e presentano un lento adattamento statico (sensori PD proporzionali- differenziali) e quindi importanti nel segnalare stimoli tattili e pressori prolungati e intensi; risultano particolarmente sensibili a forze tangenziali. In qualità di recettori articolari (meccanorecettori di stiramento) segnalano direzione e velocità di movimento nonchè la posizione dell'articolazione insieme ai meccanorecettori a rapido adattamento, sensibili all'accelerazione. Insieme ai recettori interstiziali inibiscono l'attività del sistema nervoso simpatico aumentando il tono vagale (sistema nervoso parasimpatico).
ammassi cellulari appiattiti, circondati da un rivestimento connettivale, e contengono un fitto groviglio di fibre nervose afferenti, dotate di numerose giunzioni. Essi somigliano in questo agli organi muscolo-tendinei di Golgi, con i quali hanno anche in comune la funzione principale: la misurazione della tensione nel tessuto.
 - Corpuscoli lamellari di Vater-Pacini e Paciniformi, sono corpuscoli di grandi dimensioni (fino a 4 mm di lunghezza) la cui capsula o guscio presenta fino a 60 strati di cellule appiattite circondanti una fibra nervosa mielinica di grande diametro (tipo A beta - tipo II) che perde la guaina mielinica una volta entrata nel corpuscolo.
I corpuscoli di Pacini e paciniformi si trovano immediatamente al di sotto della cute, nel tessuto adiposo ipodermico, sia nei tessuti connettivali profondi (di norma si collocano più profondamente rispetto ai corpuscoli di Ruffini), capsule articolari (strati profondi), legamenti spinali, giunzioni muscolo-tendinee. guaine di rivestimento muscolare (esterocettori cutanei e propriocettori). Essendo ad adattamento molto rapido (pochi centesimi di secondo), I corpuscoli di Vater-Pacini e paciniformi segnalano solo variazioni meccaniche molto veloci quali le vibrazioni (quelle con frequenza superiore ai 100Hz in particolare) reagendo all'inizio e al termine della sollecitazione in modo crescente (sensori di accelerazione o sensori proporzionali-crescenti PC).
- Corpuscoli lamellari di Vater-Pacini e Paciniformi, sono corpuscoli di grandi dimensioni (fino a 4 mm di lunghezza) la cui capsula o guscio presenta fino a 60 strati di cellule appiattite circondanti una fibra nervosa mielinica di grande diametro (tipo A beta - tipo II) che perde la guaina mielinica una volta entrata nel corpuscolo.
I corpuscoli di Pacini e paciniformi si trovano immediatamente al di sotto della cute, nel tessuto adiposo ipodermico, sia nei tessuti connettivali profondi (di norma si collocano più profondamente rispetto ai corpuscoli di Ruffini), capsule articolari (strati profondi), legamenti spinali, giunzioni muscolo-tendinee. guaine di rivestimento muscolare (esterocettori cutanei e propriocettori). Essendo ad adattamento molto rapido (pochi centesimi di secondo), I corpuscoli di Vater-Pacini e paciniformi segnalano solo variazioni meccaniche molto veloci quali le vibrazioni (quelle con frequenza superiore ai 100Hz in particolare) reagendo all'inizio e al termine della sollecitazione in modo crescente (sensori di accelerazione o sensori proporzionali-crescenti PC).
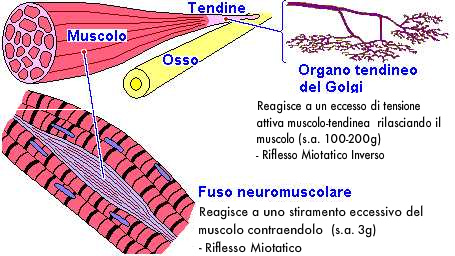 - Organi muscolo-tendinei del Golgi, sono costituiti da una capsula connettivale allungata (1 mm) contenente numerose terminazioni nervose finemente arborizzate di fibre nervose ad alta velocità di conduzione (gruppo Ib). Questi propriocettori sono situati nel tessuto connettivo denso di legamenti e capsule articolari e nelle giunzioni miotendinee (giunzione muscolo-tendinea ossia porzione del tendine che ospita le espansioni connettivali con cui terminano le fibre muscolari). Gli organi muscolo-tendinei del Golgi sono disposti in serie rispetto alle fibre fasciali e a un gruppo di fibre muscolari di riferimento (circa 20) appartenenti a diverse unità motorie.
- Organi muscolo-tendinei del Golgi, sono costituiti da una capsula connettivale allungata (1 mm) contenente numerose terminazioni nervose finemente arborizzate di fibre nervose ad alta velocità di conduzione (gruppo Ib). Questi propriocettori sono situati nel tessuto connettivo denso di legamenti e capsule articolari e nelle giunzioni miotendinee (giunzione muscolo-tendinea ossia porzione del tendine che ospita le espansioni connettivali con cui terminano le fibre muscolari). Gli organi muscolo-tendinei del Golgi sono disposti in serie rispetto alle fibre fasciali e a un gruppo di fibre muscolari di riferimento (circa 20) appartenenti a diverse unità motorie.
Gli organi del Golgi registrano la tensione muscolo-tendinea e quindi la contrazione attiva muscolare determinando, se superata la soglia di attivazione (100-200 grammi), il riflesso spinale miotatico inverso che suscita il rilassamento delle relative fibre muscolari e la contrazione di quelle del muscolo antagonista. Ricerche (Jami 1992) hanno dimostrato che la stimolazione degli organi tendinei del Golgi avviene solo se le fibre muscolari sono attivamente contratte (e non anche durante lo stiramento passivo come si credeva in passato); principio su cui si basa la tecnica PNF (Proprioceptive Neuromuscolar Facilitation).
Gli organi del Golgi registrano quindi la tensione muscolo-tendinea proteggendo le strutture tendinee, legamentose e capsulari da eccessive contrazioni muscolari ma sono altresì determinanti in ambito posturologico: la loro specifica funzione è quella di segnalare ai centri superiori la tensione media sviluppata da un selezionato gruppo di unità motorie (meccanocettori ad adattamento statico o tonico), fungendo così da meccanismo di controllo riflesso della forza di contrazione muscolare (feed-back) e di programmazione della stessa da parte dei centri motori (feed-forward). Possedendo gliorgani del Golgi una soglia di attivazione (100-200 grammi) notevolmente superiore a quelle dei fusi neuromuscolari (3 grammi), presentano di conseguenza un'azione molto meno immediata e frequente rispetto a quast'ultimi. Da notare che l'influenza dell'attività degli organi muscolotendinei del Golgi dipende molto dalle afferenze cutanee a bassa soglia e articolari.
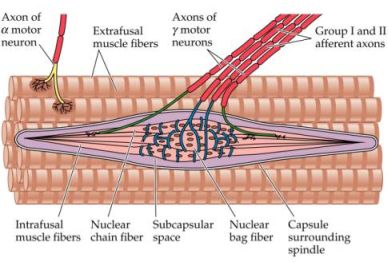 - Fusi neuromuscolari, sono recettori sensoriali diffusi nella muscolatura di tutto l'organismo, in quantità maggiore nelle vicinanze delle giunzioni muscolo-tendinee dei muscoli con funzione posturale ma anche nei muscoli dell'occhio, della mano e nei muscoli elevatori della mandibola. I fusi neuro-muscolari sono strutture formate da una capsula connettivale affusolata, dai 3 ai 10 mm di lunghezza, con all'interno 3-12 (fino a 36 nei muscoli masticatori) fibre muscolari specializzate (fibre intrafusali) poste parallelamente fra le normali fibre muscolari (extrafusali). Le fibre intrafusali sono connesse, tramite le estremità, all'endomisio delle fibre extrafusali o al tendine; la caspula che le racchiude si continua quindi con la fascia connettivale. Il fuso neuromuscolare è ricco di vasi sanguinei e nervi (più della metà delle fibre che costituiscono un nervo muscolare riguardano i fusi neuromuscolari). Sono presenti, di norma, due tipi di fibre intrafusali diverse fra loro per struttura e funzione: a sacco (in numero di 3-5 per ogni fuso con circa 100 nuclei cellulari localizzati nel rigonfiamento centrale) e a catena (8-10 per ogni fuso, con i nuclei disposti a catena sempre nella porzione centrale).
- Fusi neuromuscolari, sono recettori sensoriali diffusi nella muscolatura di tutto l'organismo, in quantità maggiore nelle vicinanze delle giunzioni muscolo-tendinee dei muscoli con funzione posturale ma anche nei muscoli dell'occhio, della mano e nei muscoli elevatori della mandibola. I fusi neuro-muscolari sono strutture formate da una capsula connettivale affusolata, dai 3 ai 10 mm di lunghezza, con all'interno 3-12 (fino a 36 nei muscoli masticatori) fibre muscolari specializzate (fibre intrafusali) poste parallelamente fra le normali fibre muscolari (extrafusali). Le fibre intrafusali sono connesse, tramite le estremità, all'endomisio delle fibre extrafusali o al tendine; la caspula che le racchiude si continua quindi con la fascia connettivale. Il fuso neuromuscolare è ricco di vasi sanguinei e nervi (più della metà delle fibre che costituiscono un nervo muscolare riguardano i fusi neuromuscolari). Sono presenti, di norma, due tipi di fibre intrafusali diverse fra loro per struttura e funzione: a sacco (in numero di 3-5 per ogni fuso con circa 100 nuclei cellulari localizzati nel rigonfiamento centrale) e a catena (8-10 per ogni fuso, con i nuclei disposti a catena sempre nella porzione centrale).
La parte recettrice del fuso è situata nella zona centrale dove, al posto dei filamenti contrattili, sono presenti le terminazione nervose dei nervi sensitivi insieme a un liquido gelatinoso (che facilita lo scorrimento delle fibre). Le fibre sensitive dei fusi neuromuscolari sono di due tipi: tipo Ia, primarie (fibre con la trasmissione più veloce dell'intero organismo), presenti una per fuso avvolte a spirale al centro di esso, e tipo II, secondarie (più lente e di diametro inferiore) situate un più in periferia e prevalentemente nelle fibre intrafusali a catena. Queste fibre nervose sensitive vengono eccitate da allungamenti anche minimi (la soglia di attivazione è circa 3 grammi) della zona centrale del fuso o dell'intero muscolo e per contrazioni delle porzioni polari delle fibre intrafusali (con la lunghezza del muscolo che non si modifica), tramite le fibre motorie gamma. Le terminazioni sensitive primarie (tipo Ia) rispondono agli stimoli in maniera dinamica (adattamento dinamico o fasico) ossia in maniera intensa solo nel momento dello stiramento della parte centrale del fuso neuromuscolare fornendo così informazioni al sistema nervoso sulla velocità dello stiramento, le terminazioni secondarie (tipo II) reagiscono agli stimoli in maniera statica (adattamento statico o tonico) ossia trasmettono segnali per tutto il tempo in cui il recettore rimane eccitato informando il sistema nervoso sull'entità dello stiramento. Oltre alle fibre sensitive Ia e II, vi è il contributo di terminazioni a fiorami, distribuite quasi esclusivamente sulle fibre intrafusali a sacco.
A ognuna delle due estremità del fuso neuromuscolare è presente una placca motrice formate da fibre nervose mieliniche motorie, più sottili rispetto a quelle muscolari, derivanti dai motoneuroni gamma delle corna anteriori del midollo spinale (classificazione delle fibre nervose). Queste fibre nervose gamma si distinguono in due tipi: fibre motorie gamma I, che innervano le fibre intrafusali a sacco e sono dinamiche (rinforzano le risposte dinamiche allo stiramento), e fibre motorie gamma II, più sottili che innervano le fibre intrafusali a sacco e a catena e sono statiche (rinforzano la reazione statica allo stiramento diminuendo o sopprimendo quella dinamica).
La complessa interazione fra le fibre nervose dei fusi neuromuscolari con i motoneuroni alfa delle fibre extrafusali costituisce il circuito o nodo gamma che determina il controllo della sensibilità dei recettori fusali (una fibra afferente Ia interagisce con più di cento motoneuroni alfa) e determina il riflesso spinale miotatico (osteotendineo, di stiramento. Tramite tale antico riflesso spinale, i fusi neuromuscolari, forniscono indicazioni sulla lunghezza del muscolo e sulla velocità di variazione della stessa lunghezza determinando il tono muscolare e fungendo da propriocettori di primaria importanza nel sistema dell'equilibrio (tonico posturale). L'importanza dei fusi neuromuscolari in ambito posturologico è pertanto enorme.
E' il sistema miofasciale (muscoli più tessuto connettivo) in realtà a rappresentare il più vasto organo sensorio del nostro organismo, è da esso infatti che il sistema nervoso centrale riceve in massima parte nervi afferenti (sensitivi). La presenza di meccanocettori, in grado di comportare effetti a livello locale e generale, è stata abbondantemente riscontrata nella fascia connettivale fin nei legamenti viscerali e nella dura madre cefalica e spinale (sacco durale).
Fra gli esterocettori cutanei e i propriocettori quelli della pianta del piede svolgono senz'altro il ruolo di protagonisti come recettori tattili. La pianta del piede è ricca di esterocettori meccanocettori sensibili alla pressione (baropressori) ad alta sensibilità (0,3 grammi), che
forniscono informazioni sulle oscillazioni corporee, e di recettori particolarmente sensibili alla trazione della cute, che quindi informano sulla direzione e sulla velocità del movimento corporeo. Essendo le informazioni dei recettori podalici le uniche a derivare direttamente da un riferimento fisso quale è il suolo, consentono di situare il corpo rispetto all'ambiente circostante. Pertanto il riflesso plantare, legato alle stimolazioni cutanee della pianta del piede, è in grado di attivare e modulare riflessi spinali molto complessi con funzioni posturali di notevole importanza (come evidenzia la rappresentazione dell'homunculus, l'area cortico-cerebrale corrispondente al piede "organo di senso" è maggiore di quella del piede "organo di moto").
In postura eretta quindi un'estesa catena propriocettiva (formata da esterocettori e propriocettori plantari, propriocettori rachidei ed enterocettori) collega, tramite uno specifico sistema di trasmissione, i recettori cefalici (occhi e apparato vestibolare, apparato stomatognatico) al recettore di riferimento fisso costituito dai piedi. Ciò consente una precisa e costante codificazione centrale, tramite il sistema tonico posturale, delle informazioni spazio-temporali cefaliche, ovvero derivanti in particolare da apparato vestibolare e vista, nonchè l'attuazione dei riflessi spinali (riflessi propriocettivi) di fondamentale importanza nell'esecuzione della gran parte dei gesti motori.
E' su tali elementi che la posturologia intende agire, tramite il programma di rieducazione posturale, una volta escluse o risolte problematiche di tipo vestibolare, visivo, neurologico ecc. Da tutto ciò si può già comprendere l'enorme influenza che può avere, in ambito posturologico, un determinato terreno e/o plantare ergonomico e/o bite.
In ambito rieducativo, occorre sottolineare come le condizioni di vita attuali (habitat artificiale con superfici quali terreno piano, sedie, scrivanie, stress col conseguente aumento dei ritmi cerebrali, sedentarietà ecc.) del tutto sfavorevoli alla biomeccanica e fisiologia umana, facilitino la dimunizione della propriocettività (dispercezione corporea) e delle abilità motorie oltre che evidentemente comportare disagi posturali, con tutto ciò che ne consegue sia a livello fisico che psichico. Da cui l'importanza vitale dell'esercizio fisico sia in campo preventivo che terapeutico e della rieducazione posturale.
Infine, traumi di vario genere sono in grado di danneggiare la struttura e la funzionalità dei recettori sensoriali alterando la propriocettività (riflessi propriocettori) e quindi la coordinazione motoria. In tali casi si rende necessaria la rieducazione motoria e, in particolare, la ginnastica propriocettiva.
Esistono varie classificazioni dei recettori sensoriali nervosi
Dal punto di vista strutturale, i recettori sensoriali possono essere più o meno e complessi e vengono distinti in tre tipi:
- Recettori tipo I, espansioni nervose formate da semplici terminazioni nervose di un singolo neurone afferente, detto neurone sensoriale primario, non mielinizzate o libere (come le terminazioni dolorifiche e tattili) o incapsulate da un rivestimento di tessuto connettivo (ad esempio recettori di pressione).
- Recettori tipo II, cellule con caratteristiche intermedie tra l'epiteliali e le nervose (cellule sensoriali), raggruppate a formare veri e propri organi di senso (ad esempio le cellule della retina per la vista e dell'organo del Corti per l'udito) e connesse, tramite sinapsi, con la fibra sensitiva (neurone primario).
- Recettori tipo III, rappresentati nell'uomo solo dai fotorecettori della retina in quanto collegati alla fibra afferente tramite un interneurone.
- tipo Ia, provenienti dalle terminazioni anulospirali dei fusi neuromuscolari, corrispondono al tipo A alfa della classificazione generale delle fibre nervose (diametro medio 17 µm);
- tipo Ib, derivano dagli organi muscolo-tendinei del Golgi, corrispondono anch'esse al tipo A alfa della classifica generale (diametro medio 16 µm);
- tipo II, provenienti dalla maggior parte dei meccanorecettori per la sensibilità cutanea fine (esterocettori cutanei), e dalle terminazioni secondarie e a fiorame dei fusi neuromuscolari, corrispondono alle fibre nervose A beta e gamma della classificazione generale (circa 8 µm di diametro);
- tipo III, relative ai recettori intersiziali, responsabili della sensibilità tattile grossolana, della termica e delle sensazioni dolorifiche puntiformi, corrispondono alle fibre A delta della classifica generale (circa 3 µm di diametro);
- tipo IV, riguardano anch'esse i recettori intersiziali, amieliniche, trasportano impulsi della sensibilità dolorifica, termica, del prurito, tattile grossolana, appartengono al tipo C della classificazione generale (diametro 0,5-2 µm).
In base alla localizzazione i recettori sensoriali vengono classificati in:
- Esterocettori, sensibili a stimoli che si originano perlopiù all'esterno dell'organismo, in sua prossimità (esterocettori cutanei) o a distanza (telecettori responsabili ad esempio di vista, udito e olfatto);
- Interocettori, sensibili a stimoli provenienti perlopiù dall'interno dell'organismo, suddivisi, a loro volta, in propriocettori, riguardanti muscoli, fascia connettivale e articolazioni (ossia fibre muscolari, tendini, legamenti, capsule articolari) ed enterocettori o propriocettori viscerali, relativi agli altri organi e sistemi dell'organismo (fra cui i recettori vestibolari, retinici e determinati barocettori renali risultano di notevole importanza riguardo l'sistema dell'equilibrio e la postura).
In base alla specializzazione, ossia al tipo di stimoli rilevati con più bassa soglia di atttivazione, è possibile distinguere fondamentalmente cinque tipi di recettori:
- Meccanocettori (meccanorecettori), segnalano le deformazioni del recettore stesso o delle cellule ad esso adiacenti;
- Termocettori (termorecettori), terminazioni nervose libere, a lento adattamento, che segnalano variazioni di temperatura sia interna che esterna all'organismo (sensibili o al caldo, calorecettori, o al freddo, frigorecettori);
- Nocicettori, recettori dolorifici, costituiti perlopiù da terminazioni nervose libere, che segnalano un danno tessutale fisico e/o chimico;
- Recettori elettromagnetici, segnalano gli stimoli luminosi a livello della retina (fotorecettori);
- Chemocettori (chemiocettori), segnalano stimoli di natura chimica (es. quelli gustativi della bocca, olfattivi del naso, quelli che segnalano la concentrazione di ossigeno nel sangue, il ph di sangue e liquor cerebrospinale, l'osmolarità dei liquidi corporei ecc.).
- Recettori fasici o dinamici o di velocità, si attivano solo quando l'intensità dello stimolo cambia inviando un numero di impulsi proporzionale alla velocità della variazione per poi, tramite il fenomeno di rapido adattamento, divenire presto silenti una volta che lo stimolo rimane costante. Questo tipo di recettori consente al sistema nervoso centrale di conoscere la velocità con cui sta avvenendo un certo cambiamento nel corpo, consentendogli così di effettuare previsioni e quindi opportuni adeguamenti anticipatori (feed-forward). Le terminazioni sensitive primarie (tipo Ia) dei fusi neuromuscolari (propriocettori) e i corpuscoli di Pacini (esterocettori cutanei meccanocettori) sono importanti esempi di recettori di questo tipo.
- Recettori tonici o statici, i quali, in presenza di uno stimolo costante e prolungato, continuano a trasmettere fino alla cessazione dello stimolo o almeno per molti minuti od ore (fenomeno di lento adattamento). Esempi di questo tipo di recettori sono gli organi muscolo-tendinei del Golgi e le terminazioni nervose sensitive secondarie (tipo II) dei fusi neuromuscolari (propriocettori), i corpuscoli di Ruffini e i dischi di Merkel (esterocettori cutanei sensibili al tatto).
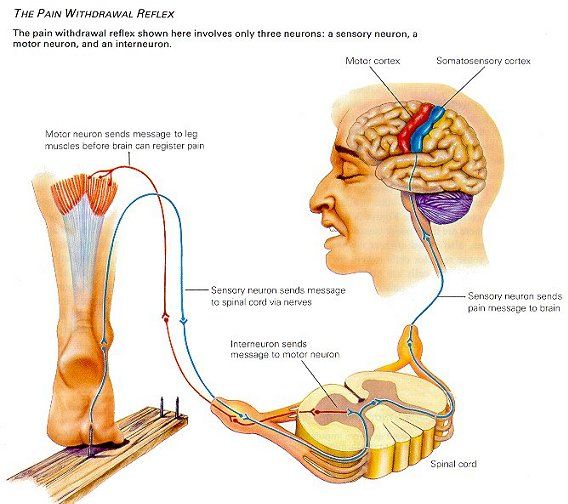 Il neurone sensitivo primario (neurone primario) porta il segnale fino al midollo spinale (corna posteriori) dove, nella maggior parte dei casi, contrae sinapsi con interneuroni o neuroni sensitivi secondari (ogni fibra nervosa in arrivo si ramifica centinaia o migliaia di volte contraendo sinapsi con i dendriti o i corpi cellulari dei neuroni di un determinato pool neuronale, definendo così il proprio "campo di stimolazione"). Questi ultimi, a loro volta, possono contrarre sinapsi ancora a livello del midollo spinale o raggiungere il talamo (tranne l'olfatto che prende direttamente contatto con l'encefalo), dove si connettono con i neuroni sensitivi terziari che prendono ulteriori contatti col sistema nervoso centrale. Da sottolineare il ruolo dei nuclei vestibolari a cui arrivano le informazioni di tutti i recettori sensoriali posturali (recettori vestibolari, esterocettori cutanei, propriocettori ed enterocettori ed esterocettori visivi) e vengono elaborate, assieme alla sostanza reticolare e sotto il controllo del cervelletto, oltre che della corteccia cerebrale, consentendo così al sistema dell'equilibrio (sistema tonico posturale) di svolgere il suo compito, ossia di garantire il corretto assetto posturale sia statico che dinamico.
Il neurone sensitivo primario (neurone primario) porta il segnale fino al midollo spinale (corna posteriori) dove, nella maggior parte dei casi, contrae sinapsi con interneuroni o neuroni sensitivi secondari (ogni fibra nervosa in arrivo si ramifica centinaia o migliaia di volte contraendo sinapsi con i dendriti o i corpi cellulari dei neuroni di un determinato pool neuronale, definendo così il proprio "campo di stimolazione"). Questi ultimi, a loro volta, possono contrarre sinapsi ancora a livello del midollo spinale o raggiungere il talamo (tranne l'olfatto che prende direttamente contatto con l'encefalo), dove si connettono con i neuroni sensitivi terziari che prendono ulteriori contatti col sistema nervoso centrale. Da sottolineare il ruolo dei nuclei vestibolari a cui arrivano le informazioni di tutti i recettori sensoriali posturali (recettori vestibolari, esterocettori cutanei, propriocettori ed enterocettori ed esterocettori visivi) e vengono elaborate, assieme alla sostanza reticolare e sotto il controllo del cervelletto, oltre che della corteccia cerebrale, consentendo così al sistema dell'equilibrio (sistema tonico posturale) di svolgere il suo compito, ossia di garantire il corretto assetto posturale sia statico che dinamico.
La sensibilità cosciente ai segnali in partenza dai recettori nervosi si definisce epicritica, ossia fine e localizzata, e ne è responsabile la corteccia cerebrale, oppure protopatica, ossia diffusa (non ben localizzata) e "grossolana" (difficilmente definibile), ed è sotto la gestione del talamo.
Il tipo di sensazioni (modalità sensoriale) o risposte evocate dalla stimolazione di un dato recettore dipende esclusivamente dalla regione del sistema nervoso centrale attivata dalla relativa via sensitiva. Indipendentemente dalla natura dello stimolo e dal punto di applicazione di esso lungo una via sensitiva, le sensazioni relative saranno le stesse (ad esempio se viene eccitata una fibra nervosa dolorifica, si avverte dolore indipendentemente dalla natura dello stimolo che lo ha provocato) e verranno riferite di norma alla sede del recettore (e non nella zona di stimolazione); come nel caso del "arto fantasma" in cui talvolta pur avendo un arto amputato si lamentano dolori proveniente dall'arto assente. Spesso però, in caso di problemi muscolo-scheletrici o viscerali, è possibile avvertire dolore, a livello cutaneo, in una sede distante dall'origine di esso (dolore riferito). La zona cutanea di riferimento corrisponde al dermatomero le cui fibre afferenti (sensoriali) entrano nel midollo spinale tramite la medesima radice posteriore in cui decorrono anche fibre sensoriali viscerali o muscolari.
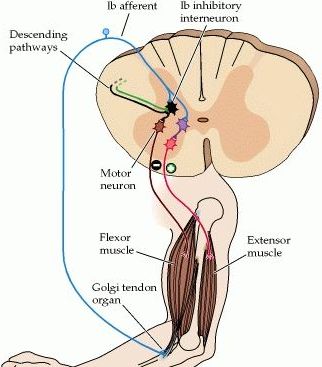 Nel riflesso spinale miotatico inverso, le terminazioni nervose amieliniche degli organi muscolo-tendinei del Golgi subiscono, durante la contrazione attiva del muscolo, una deformazione meccanica che innesca un impulso nervoso che giunge al midollo spinale, lungo fibre nervose ad alta velocità di conduzione del gruppo Ib (classificazione fibre nervose), inibendo i motoneuroni alfa tramite l'intermediazione di un solo interneurone spinale inibitorio (connessione disinaptica ossia tramite due sinapsi) mentre, in contemporanea, un interneurone facilitatorio stimola la contrazione dei muscoli antagonisti (attivazione antagonista). La soglia di attivazione del riflesso miotatico inverso è di 100-200 grammi (la soglia di attivazione del riflesso miotatico è di 3 grammi).
Nel riflesso spinale miotatico inverso, le terminazioni nervose amieliniche degli organi muscolo-tendinei del Golgi subiscono, durante la contrazione attiva del muscolo, una deformazione meccanica che innesca un impulso nervoso che giunge al midollo spinale, lungo fibre nervose ad alta velocità di conduzione del gruppo Ib (classificazione fibre nervose), inibendo i motoneuroni alfa tramite l'intermediazione di un solo interneurone spinale inibitorio (connessione disinaptica ossia tramite due sinapsi) mentre, in contemporanea, un interneurone facilitatorio stimola la contrazione dei muscoli antagonisti (attivazione antagonista). La soglia di attivazione del riflesso miotatico inverso è di 100-200 grammi (la soglia di attivazione del riflesso miotatico è di 3 grammi).
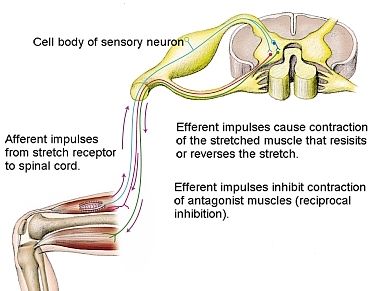 Gli impulsi provenienti dai fusi neuromuscolari, durante l’allungamento attivo o passivo del muscolo, raggiungono il midollo spinale ed eccitano i motoneuroni alfa dello stesso muscolo e dei muscoli agonisti tramite una trasmissione monosinaptica (tramite una sinapsi), ciò provoca il riflesso spinale miotatico o di stiramento o riflesso osteotendineo (ROT), ossia una contrazione riflessa che tende a riportare il muscolo alla sua lunghezza originaria. Le afferenze nervose secondarie, tipo II (classificazione fibre nervose), accanto ad una funzione monosinaptica, esercitano complessi effetti polisinaptici di tipo eccitatorio sui muscoli flessori e di tipo inibitorio sui muscoli estensori. Come tutti i riflessi spinali, il riflesso miotatico agisce in maniera gerarchica ed è sottoposto, in maniera parallela, al controllo/gestione dei centri superiori. Essenso la soglia di attivazione del riflesso miotatico molto bassa (3 grammi), l'attività dei fusi neuromuscolari è praticamemente permanente e da loro dipende il tono muscolare con grosse infleunze sulla postura. Tale riflesso avendo il compito di mantenere costante la lunghezza di un muscolo viene sempre utilizzato per stabilizzare le diverse posture; il riflesso di stiramento muscolare o riflesso miotatico o osteotendineo (ROT) è infatti il più antico e il primo riflesso a esserne implicato.
Gli impulsi provenienti dai fusi neuromuscolari, durante l’allungamento attivo o passivo del muscolo, raggiungono il midollo spinale ed eccitano i motoneuroni alfa dello stesso muscolo e dei muscoli agonisti tramite una trasmissione monosinaptica (tramite una sinapsi), ciò provoca il riflesso spinale miotatico o di stiramento o riflesso osteotendineo (ROT), ossia una contrazione riflessa che tende a riportare il muscolo alla sua lunghezza originaria. Le afferenze nervose secondarie, tipo II (classificazione fibre nervose), accanto ad una funzione monosinaptica, esercitano complessi effetti polisinaptici di tipo eccitatorio sui muscoli flessori e di tipo inibitorio sui muscoli estensori. Come tutti i riflessi spinali, il riflesso miotatico agisce in maniera gerarchica ed è sottoposto, in maniera parallela, al controllo/gestione dei centri superiori. Essenso la soglia di attivazione del riflesso miotatico molto bassa (3 grammi), l'attività dei fusi neuromuscolari è praticamemente permanente e da loro dipende il tono muscolare con grosse infleunze sulla postura. Tale riflesso avendo il compito di mantenere costante la lunghezza di un muscolo viene sempre utilizzato per stabilizzare le diverse posture; il riflesso di stiramento muscolare o riflesso miotatico o osteotendineo (ROT) è infatti il più antico e il primo riflesso a esserne implicato.
- Postura e Benessere, Giovanni Chetta
- La Scoliosi quale atteggiamento naturale, Giovanni Chetta
- Il Sistema Connettivo, Giovanni Chetta
- Dalla MEC alla postura, Giovanni Chetta
- Piede e postura, Giovanni Chetta
- Ginnastica Posturale TIB, Giovanni Chetta
- Massaggio & Bodywork TIB, Giovanni Chetta
- Stress e Benessere, Giovanni Chetta
