Approfondimenti |
|

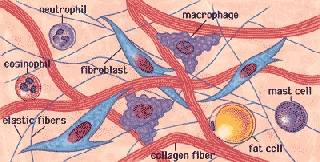 Il tessuto connettivo si sviluppa dal tessuto embrionale mesenchima caratterizzato da cellule ramificate comprese in un'abbondante sostanza intercellulare amorfa. Il mesenchima deriva dal foglietto embrionale intermedio, mesoderma, molto diffuso nel feto dove circonda gli organi in via di sviluppo compenetrandoli. Alcuni tessuti connettivi del cranio derivano direttamente dal neuroectoderma (fascia meningea). Il mesenchima, oltre a produrre tutti i tipi di tessuto connettivo, produce altri tessuti: muscolare, vasi sanguinei, epitelio e alcune ghiandole.
Il tessuto connettivo si sviluppa dal tessuto embrionale mesenchima caratterizzato da cellule ramificate comprese in un'abbondante sostanza intercellulare amorfa. Il mesenchima deriva dal foglietto embrionale intermedio, mesoderma, molto diffuso nel feto dove circonda gli organi in via di sviluppo compenetrandoli. Alcuni tessuti connettivi del cranio derivano direttamente dal neuroectoderma (fascia meningea). Il mesenchima, oltre a produrre tutti i tipi di tessuto connettivo, produce altri tessuti: muscolare, vasi sanguinei, epitelio e alcune ghiandole.
Il tessuto connettivo è morfologicamente caratterizzato da vari tipi di cellule (fibroblasti, macrofagi, mastociti, plasmacellule, leucociti, cellule indifferenziate, cellule adipose o adipociti, condrociti, osteociti, miofibroblasti ecc.) immersi in un abbondante materiale intercellulare, definito MEC (matrice extracellulare) o ECM (extracellular matrix), sintetizzato dalle stesse cellule connettivali. La MEC è composta da fibre proteiche insolubili (collagene, elastiche e reticolari) e sostanza fondamentale, erroneamente definita amorfa, colloidale, formata da complessi solubili di carboidrati, in gran parte legati a proteine, detti mucopolisaccaridi acidi, glicoproteine, proteoglicani, glucosamminoglicani o GAG (acido ialuronico, coindroitinsolfato, cheratinsolfato, eparinsolfato ecc.) e, in minor misura, da proteine specializzate, fra cui la fibronectina.
Cellule e matrice intercellulare caratterizzano vari tipi di tessuto connettivo: tessuto connettivo propriamente detto (fascia connettivale), tessuto elastico, tessuto reticolare, tessuto mucoso, tessuto endoteliale, tessuto adiposo, tessuto cartilagineo, tessuto osseo, sangue e linfa. I tessuti connettivi giocano quindi diversi importanti ruoli: strutturali, difensivi, trofici e morfogenetici organizzando e influenzando la crescita e la differenziazione dei tessuti circostanti.
Le condizioni della parte fibrosa e della sostanza fondamentale del sistema connettivale sono in parte determinate dalla genetica, in parte da fattori ambientali (nutrizione, esercizio fisico ecc.). Le fibre proteiche sono infatti in grado di modificarsi in base alle esigenze ambientali e funzionali. Del loro spettro di variabilità strutturale e funzionale ne sono esempi il tegumento, la membrana basale, la cartilagine, l'osso, i legamenti, i tendini ecc. La sostanza fondamentale varia continuamente il suo stato, divenendo più o meno viscosa (da fluida a collosa fino a solida), in base alle specifiche esigenze organiche. Riscontrabile in grandi quantità quale liquido sinoviale articolare e umor vitreo oculare, essa è in realtà presente in tutti i tessuti.
Il tessuto connettivo varia le proprie caratteristiche strutturali attraverso l'effetto piezo-elettrico: qualunque forza meccanica che crea deformazione strutturale stira i legami inter-molecolari producendo un leggero flusso elettrico (carica piezoelettrica). Questa carica può essere rilevata dalle cellule e comportare modifiche biochimiche: ad esempio, nell'osso, gli osteoclasti non possono "digerire" osso piezoelettricamente carico.
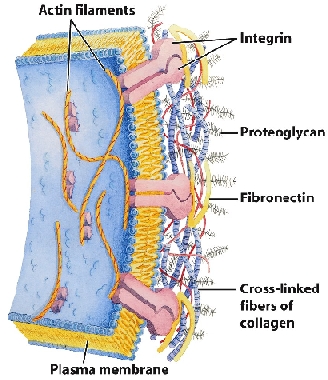 La matrice extracellulare fornisce l'ambiente chimico-fisico per le cellule che avvolge (prendendo così parte alla regolazione dell'equilibrio acido-base, del metabolismo idrosalino, dell'equilibrio elettrico e osmotico) formando una struttura a cui queste aderiscono e dentro la quale possono muoversi mantenendo un adatto ambiente ionico idratato e permeabile, attraverso il quale diffondono i metaboliti. La densità della matrice fibrosa e la viscosità della sostanza fondamentale determinano il libero flusso delle sostanze chimiche fra cellule impedendo allo stesso tempo la penetrazione di batteri e particelle inerti. Presenta cellule del sistema immunitario (leucociti, macrofagi, mastociti, plasmacellule) ed è frequentemente il luogo di svolgimento dei processi infiammatori. Esso inoltre possiede grandi capacità riparative delle zone danneggiate da infiammazioni e/o traumi riempiendone, se necessario, gli spazi.
La matrice extracellulare fornisce l'ambiente chimico-fisico per le cellule che avvolge (prendendo così parte alla regolazione dell'equilibrio acido-base, del metabolismo idrosalino, dell'equilibrio elettrico e osmotico) formando una struttura a cui queste aderiscono e dentro la quale possono muoversi mantenendo un adatto ambiente ionico idratato e permeabile, attraverso il quale diffondono i metaboliti. La densità della matrice fibrosa e la viscosità della sostanza fondamentale determinano il libero flusso delle sostanze chimiche fra cellule impedendo allo stesso tempo la penetrazione di batteri e particelle inerti. Presenta cellule del sistema immunitario (leucociti, macrofagi, mastociti, plasmacellule) ed è frequentemente il luogo di svolgimento dei processi infiammatori. Esso inoltre possiede grandi capacità riparative delle zone danneggiate da infiammazioni e/o traumi riempiendone, se necessario, gli spazi.
Nel tessuto adiposo, che costituisce un tipo di tessuto connettivo, si accumulano i lipidi, importanti riserve nutritizie mentre nel tessuto connettivo lasso si conserva acqua ed elettroliti (grazie al suo alto contenuto di mucopolisacaridi acidi) e circa 1/3 delle proteine plasmatiche totali sono nel compartimento intercellulare del tessuto connettivo
Combinando una piccola varietà di fibre all'interno di una matrice che varia da fluida a collosa a solida, le cellule connettivali rispondono alle esigenze di flessibilità e stabilità, diffusione e barriera.
 Dal punto di vista meccanico la MEC (matrice extracellulare) si è sviluppata per distribuire le tensioni del movimento e della gravità mantenendo contemporaneamente la forma dei diversi componenti del corpo tramite tutta la gamma di possibilità che va dalla rigidità di una struttura a compressione continua all'elasticità di una struttura a tensegrità. Nella struttura di tensegrità le parti in compressione (le ossa) spingono in fuori contro le parti in trazione (miofascia) che spingono verso l'interno. Questo tipo di strutture presentano una stabilità più elastica rispetto a quelle a compressione continua e diventano tanto più stabili quanto più vengono caricate. Tutti gli elementi interconnessi di una struttura a tensegrità si ridispongono in risposta a una tensione locale.
Lo stesso scheletro è in realtà solo apparentemente una struttura a compressione continua in quanto le ossa poggiano su superfici scivolose (cartilagini articolari) e senza il sostegno miofasciale non sono in grado di sostenersi. Quindi variare la tensione dei tessuti molli significa variare la disposizione delle ossa e la minima variazione strutturale di un "angolo" organico si trasmette meccanicamente e piezolettricamente, tramite la rete di tensegrità, su tutte le restanti parti corporee.
Dal punto di vista meccanico la MEC (matrice extracellulare) si è sviluppata per distribuire le tensioni del movimento e della gravità mantenendo contemporaneamente la forma dei diversi componenti del corpo tramite tutta la gamma di possibilità che va dalla rigidità di una struttura a compressione continua all'elasticità di una struttura a tensegrità. Nella struttura di tensegrità le parti in compressione (le ossa) spingono in fuori contro le parti in trazione (miofascia) che spingono verso l'interno. Questo tipo di strutture presentano una stabilità più elastica rispetto a quelle a compressione continua e diventano tanto più stabili quanto più vengono caricate. Tutti gli elementi interconnessi di una struttura a tensegrità si ridispongono in risposta a una tensione locale.
Lo stesso scheletro è in realtà solo apparentemente una struttura a compressione continua in quanto le ossa poggiano su superfici scivolose (cartilagini articolari) e senza il sostegno miofasciale non sono in grado di sostenersi. Quindi variare la tensione dei tessuti molli significa variare la disposizione delle ossa e la minima variazione strutturale di un "angolo" organico si trasmette meccanicamente e piezolettricamente, tramite la rete di tensegrità, su tutte le restanti parti corporee.
Fascia connettivale e muscoli costituiscono, anatomicamente e funzionalmente, il sistema miofasciale il cui tessuto connettivo è sede di numerosissimi recettori sensoriali, inclusi gli esterocettori e i propriocettori nervosi e struttura i muscoli, anatomicamente e funzionalmente, in catene miofasciali, assumendo pertanto un ruolo fondamentale all'interno del sistema dell'equilibrio e della postura; è nella rete connettivale che registriamo postura e pattern di movimento tramite la comunicazione meccanica connettivale, la quale incide in ciò più dei meccanismi riflessi dei fusi neuromuscolari e degli organi tendinei del Golgi (organi di senso propriocettivi attraverso cui il sistema nervoso si informa su ciò che accade nella rete miofasciale). E' infatti il sistema miofasciale in realtà il più vasto organo sensorio del nostro organismo, è da esso che il sistema nervoso centrale riceve in massima parte nervi afferenti (sensitivi). La presenza di meccanocettori, in particolare i recettori interstiziali, in grado di comportare effetti a livello locale e generale, è stata abbondantemente riscontrata nella fascia connettivale fin nei legamenti viscerali e nella dura madre cefalica e spinale (sacco durale).
 Nel sistema miofasciale (muscolo-fasciale) del nostro corpo, ciascun muscolo, organo viscerale, vaso (sanguineo e linfatico), nervo, osso, articolazione ecc. è rinchiuso nel proprio involucro fasciale. Tali involucri, a loro volta, formano un reticolo
ubiquitario di tensegrità, che avvolge e, al tempo stesso, sostiene e collega tutte le unità funzionali del corpo. Infine, questi robusti strati elastici formano anche uno strato superficiale, che funge da contenitore e da sostegno frenante per tutto il corpo: le fasce superficiali poste sotto la pelle.
Nel sistema miofasciale (muscolo-fasciale) del nostro corpo, ciascun muscolo, organo viscerale, vaso (sanguineo e linfatico), nervo, osso, articolazione ecc. è rinchiuso nel proprio involucro fasciale. Tali involucri, a loro volta, formano un reticolo
ubiquitario di tensegrità, che avvolge e, al tempo stesso, sostiene e collega tutte le unità funzionali del corpo. Infine, questi robusti strati elastici formano anche uno strato superficiale, che funge da contenitore e da sostegno frenante per tutto il corpo: le fasce superficiali poste sotto la pelle.
Il muscolo è tenuto in sede tramite lamine connettivali (aponeurosi o aponevrosi) ed è racchiuso nelle fasce come la polpa di un'arancia lo è nelle pareti cellulari che la suddividono (epimisio, perimisio ed endomisio). "Ostruzioni" locali, come ad es. le aderenze fasciali, possono derivare da sforzi eccessivi o mancanza di esercizio, traumi, malattie infiammatorie, cicatrici aderenziali ecc. In presenza di aderenze fasciali nei diversi strati fasciali, si ha un aumentato attrito interno che contrasta il movimento e, in particolare, l'allungamento muscolare. Ciò crea un trazionamento delle strutture adiacenti che concorre all'affaticamento e alle tensioni generali.
Inoltre, un muscolo che lavora persistentemente in accorciamento, oltre a modificare in qualità e quantità (in base alle forze su essa agenti e agli spazi disponibili) la porzione connettivale, può diminuire il numero di sarcomeri (al contrario, un muscolo che lavora in allungamento tende ad aumentare la parte connettivale e il numero di sarcomeri); si ha così la formazione di un muscolo retratto. L'eliminazione di tali impedimenti e quindi il ripristino della corretto flusso consente alle cellule interessate di passare da un metabolismo di sopravvivenza a quello fisiologico specifico.
La scoperta della presenza di cellule del tessuto connettivo interposte alle fibre collagene fasciali con capacità contrattili simili alla muscolatura liscia, denominate miofibroblasti, ha dimostrato la capacità del tessuto connettivo di contrarsi in determinate situazioni.
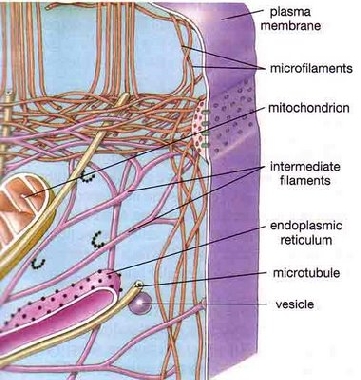 L'avanzamento tecnico della microscopia elettronica ha dimostrato che la cellula è tutt'altro che un sacco membranoso contenente una soluzione di molecole, come si credeva in passato. La cellula è in realtà piena di filamenti, tubicini, fibre e trabecole formanti una struttura definita matrice citoplasmatica o citoscheletro. Vi è ben poco spazio a disposizione per consentire la casuale diffusione di molecole, inoltre pochissima acqua è presente allo stato libero essendo quasi totalmente in stato di solvazione, così come accade per il tessuto connettivo.
L'avanzamento tecnico della microscopia elettronica ha dimostrato che la cellula è tutt'altro che un sacco membranoso contenente una soluzione di molecole, come si credeva in passato. La cellula è in realtà piena di filamenti, tubicini, fibre e trabecole formanti una struttura definita matrice citoplasmatica o citoscheletro. Vi è ben poco spazio a disposizione per consentire la casuale diffusione di molecole, inoltre pochissima acqua è presente allo stato libero essendo quasi totalmente in stato di solvazione, così come accade per il tessuto connettivo.
Il citoscheletro è in massima parte costituito dai microfilamenti di actina, una proteina globulare, e dai microtubuli di tubulina, una proteina tubulare. Microtubuli e microfilamenti si formano e si disgregano spontaneamente al presentarsi di particolari condizioni ambientali (es.: presenza di Ca2+ e Mg2+).
Già all'inizio degli anni ottanta si era compreso il ruolo del citoscheletro nel sostegno della cellula, nel consentire gli spostamenti della stessa cellula e delle vescicole e la sua implicazione nei processi di divisione cellulare. Inoltre, si era evidenziato come la matrice extracellulare risulta legata al sistema del citoscheletro così da tenere assieme il nostro corpo. Oggi sappiamo che tali legami incidono su processi fisiologici quali lo sviluppo embrionale, la coagulazione del sangue, la guarigione delle ferite ecc.
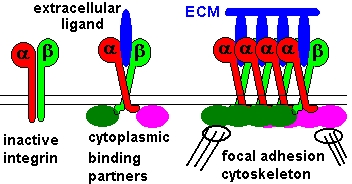 Vi sono connessioni meccanicamente mutuamente attive tra l'interno della cellula e la MEC (matrice extracellulare). Ciò cancella del tutto l'idea che le cellule fluttuino indipendenti fra loro all'interno di una sostanza "amorfa". Il doppio involucro della membrana fosfolipidica cellulare, infatti, oltre a essere costellato, sia esternamente che internamente, da chemiorecettori (proteine globulari con siti recettoriali per specifici agenti chimici capaci di modificare l'attività della cellula), presenta alcune glicoproteine di membrana a struttura bicatenaria, definite integrine, che fungono da meccanocettori. Esse interagendo con le proteine della matrice extracellulare, fattori del Complemento, ecc., trasmettono trazioni e spinte meccaniche dalla matrice fibrosa connettiva extracellulare all'interno della cellula e viceversa.
Le integrine appaiono virtualmente su ogni tipo di cellule animali e sembrano essere i principali recettori mediante i quali le cellule aderiscono alla matrice extracellulare e sono in grado di mediare importanti eventi di adesione cellula-cellula. Inoltre è stata dimostrata la loro capacità di trasdurre, selettivamente e in maniera modulabile, segnali all'interno e all'esterno della cellula in un'ampia varietà di tipi cellulari, anche in sinergismo con altri sistemi recettoriali. Le integrine sono quindi versatili molecole che giocano un ruolo chiave in vari processi cellulari sia durante lo sviluppo sia nell'organismo adulto: adesione e migrazione cellulare, crescita e divisione cellulare, sopravvivenza, apoptosi e differenziazione cellulare, sostegno al sistema immunitario ecc. Varie malattie genetiche umane dimostrano l'importanza di queste molecole in vari processi fisiologici e patologici. Questa conessione specializzata (strutturale-funzionale) MEC-citoscheletro viene definita adesione focale.
La meccanica delle connessioni tra la matrice extracellulare e quella intracellulare è raggiunta attraverso una numerosa serie di legami deboli (non covalenti) e indiretti, tramite proteine "armatura" (talina, paxillina, alpha-actinina ecc.), che si connettono o disconnettono in maniera rapida (una specie di effetto velcro). Le cellule sono quindi tra loro connesse attraverso una matrice che comunica con loro attraverso attivi legami deboli secondo una geometria di tensegrità che varia costantemente in base all'attività della cellula e del corpo e alla condizione della matrice stessa.
La connessione della cellula alla matrice extracellulare (MEC) è un requisito base per formare un organismo pluricellulare. Essa rende la cellula in grado di resistere alle forze tiranti senza essere sbalzata fuori dalla MEC. Inoltre, le integrine rappresentano i piedi che consentono alla cellula la migrazione nel substrato extracellulare.
Attraverso il citoscheletro la comunicazione meccanica raggiunge anche il nucleo. Queste connessioni agiscono cambiando la forma della cellula e quindi le proprietà fisiologiche.
Vi sono connessioni meccanicamente mutuamente attive tra l'interno della cellula e la MEC (matrice extracellulare). Ciò cancella del tutto l'idea che le cellule fluttuino indipendenti fra loro all'interno di una sostanza "amorfa". Il doppio involucro della membrana fosfolipidica cellulare, infatti, oltre a essere costellato, sia esternamente che internamente, da chemiorecettori (proteine globulari con siti recettoriali per specifici agenti chimici capaci di modificare l'attività della cellula), presenta alcune glicoproteine di membrana a struttura bicatenaria, definite integrine, che fungono da meccanocettori. Esse interagendo con le proteine della matrice extracellulare, fattori del Complemento, ecc., trasmettono trazioni e spinte meccaniche dalla matrice fibrosa connettiva extracellulare all'interno della cellula e viceversa.
Le integrine appaiono virtualmente su ogni tipo di cellule animali e sembrano essere i principali recettori mediante i quali le cellule aderiscono alla matrice extracellulare e sono in grado di mediare importanti eventi di adesione cellula-cellula. Inoltre è stata dimostrata la loro capacità di trasdurre, selettivamente e in maniera modulabile, segnali all'interno e all'esterno della cellula in un'ampia varietà di tipi cellulari, anche in sinergismo con altri sistemi recettoriali. Le integrine sono quindi versatili molecole che giocano un ruolo chiave in vari processi cellulari sia durante lo sviluppo sia nell'organismo adulto: adesione e migrazione cellulare, crescita e divisione cellulare, sopravvivenza, apoptosi e differenziazione cellulare, sostegno al sistema immunitario ecc. Varie malattie genetiche umane dimostrano l'importanza di queste molecole in vari processi fisiologici e patologici. Questa conessione specializzata (strutturale-funzionale) MEC-citoscheletro viene definita adesione focale.
La meccanica delle connessioni tra la matrice extracellulare e quella intracellulare è raggiunta attraverso una numerosa serie di legami deboli (non covalenti) e indiretti, tramite proteine "armatura" (talina, paxillina, alpha-actinina ecc.), che si connettono o disconnettono in maniera rapida (una specie di effetto velcro). Le cellule sono quindi tra loro connesse attraverso una matrice che comunica con loro attraverso attivi legami deboli secondo una geometria di tensegrità che varia costantemente in base all'attività della cellula e del corpo e alla condizione della matrice stessa.
La connessione della cellula alla matrice extracellulare (MEC) è un requisito base per formare un organismo pluricellulare. Essa rende la cellula in grado di resistere alle forze tiranti senza essere sbalzata fuori dalla MEC. Inoltre, le integrine rappresentano i piedi che consentono alla cellula la migrazione nel substrato extracellulare.
Attraverso il citoscheletro la comunicazione meccanica raggiunge anche il nucleo. Queste connessioni agiscono cambiando la forma della cellula e quindi le proprietà fisiologiche.
Gli studi effettuati da Ingber D. e pubblicati sulla rivista "Scientific American" nel 1998, infatti, hanno dimostrato che semplicemente modificando la forma cellulare è possibile innescare diversi processi genetici. Forzando le cellule viventi a prendere diverse forme piazzandole su "isole adesive" composte da matrice extracellulare, risultò che le cellule piatte, stirate, avevano più probabilità di dividersi, interpretando tale stato come bisogno di fornire più cellule per riempire lo spazio circostante (come ad es nel caso di ferite), quella rotondeggianti, a cui veniva impedito di propagarsi comprimendole, attivavano un programma di morte per apoptosi (morte funzionale programmata) per evitare un sovraffollamento in grado di generare tumori. Quando invece le cellule non erano nè troppo espanse nè troppo compresse eseguivano specifiche fisiologiche attività in base alla loro provenienza e differenziazione (le cellule capillari formavano tubi capillari cavi, le epatiche secernevano le tipiche proteine fornite dal fegato al sangue ecc.),
La maggior parte degli studi sul cancro si concentra sui segnali chimici ma i legami fra il microambiente tissutale e l'oncogenesi potrebbero permettere l'identificazione di nuovi target terapeutici; i tessuti tumorali sono più rigidi dei tessuti normali e la palpazione di una massa rigida è talvolta un utile metodo per individuarne la presenza. Uno studio, incentrato sulle integrine e pubblicato sulla rivista "Cancer Cell" nel settembre 2005 (Matthew J. Paszek, et al., “Tensional homeostasis and the malignant phenotype”), ha messo in luce un legame fra la rigidità dei tessuti e la formazione dei tumori evidenziando come le forze meccaniche possono regolare il comportamento cellulare influenzando i segnali molecolari che governano la diffusione delle cellule cancerose. I ricercatori hanno esaminato cellule cancerose in sviluppo all'interno di un sistema gelatinoso tridimensionale, nel quale la rigidità poteva essere controllata accuratamente. Essi hanno scoperto che anche un leggero aumento di durezza della matrice extracellulare circostante perturba l'architettura dei tessuti e ne favorisce la crescita promuovendo l'adesione focale e l'attivazione dei fattori di crescita. Mentre, un calo di attività Rho o ERK (enzimi costituenti fattori oncogeni in quanto frequentemente coinvolti nel processo di metastatizzazione), nelle cellule cancerose, è stato associato a un susseguente declino di adesione focale e all'inversione delle modifiche morfologiche. Tuttavia, la relazione fra la rigidità tissutale e il comportamento delle cellule tumorali non è ancora del tutto chiara.
Di seguito un video sulla struttura e vita interna della cellula
La struttura totale costituita da matrice extracellulare (MEC), recettori di membrana (integrine) e matrice intracellulare costituisce il nostro corpo spaziale (da cui il concetto di "medicina e salute spaziale"). Ci troviamo di fronte a un vero e proprio network sopramolecolare continuo e dinamico che si estende in ogni angolo e spazio corporeo composto da una matrice nucleare interna a una matrice cellulare immersa in una matrice extracellulare. Il network connettivale è un continuum che connette sia strutturalmente che funzionalmente dando forma e consentendo la vita del nostro organismo.
A differenza dei networks formati dal sistema nervoso, da quello endocrino e da quello immunitario, il sistema connettivo presenta un metodo forse apparentemente più arcaico ma non certo meno importante di comunicazione: quella meccanica. Esso "semplicemente" tira e spinge comunicando così da fibra a fibra, da cellula a cellula e da ambiente interno ed esterno alla cellula e viceversa, tramite la trama fibrosa, la sostanza fondamentale e sofisticati sistemi di transduzione del segnale meccanico. Questo tipo di comunicazione avviene al di fuori del nostro stato di coscienza ed è stato fin'ora studiato molto meno della comunicazione nervosa e circolatoria.
Occorre inoltre considerare che il sistema connettivo, oltre a rappresentare esso stesso un network indispensabile per lo sviluppo e la vita di tutti i componenti dell'organismo, costituisce il substrato integrato che consente agli altri networks (nervoso, endocrino, e immunitario) di esistere, agire e comunicare fra loro. In altre parole, esso è al contempo un sistema di comunicazione integrato con gli altri nonchè la struttura fisicamente e funzionalmente portante di essi e quindi della rete globale organica.
Al contempo, i sistemi endocrino, immunitario e nervoso sono in grado di comportare profonde modifiche del sistema connettivo come, ad esempio, nei processi di cicatrizzazione e infiammatori o, semplicemente, considerando le modifiche fasciali comportate dai muscoli tramite il sistema nervoso (possiamo considerare in realtà i muscoli come un'unica gelatina, che modifica rapidamente il proprio stato in risposta a uno stimolo nervoso, contenuta all'interno di 650 tasche connettivali).
Ulteriore fattore in grado di incidere in maniera preponderante sul sistema connettivo è l'alimentazione (lo scorbuto, ad esempio, è una patologia caratterizzata da un'estesa degenerazione del tessuto connettivo dovuta a carenza di vitamina C, in assenza della quale i fibroblasti cessano di sintetizzare il collagene).
In circa 4 miliardi di anni vita su questo pianeta, gli esseri umani si sono evoluti quali aggregati di circa 6 trilioni di quattro diversi tipi di cellule disperse all'interno di un elemento fluido: cellule nervose, specializzate nella conduzione, muscolari specializzate nella contrazione, epiteliali specializzate nella secrezione (enzimi, ormoni ecc.) e connettivali. Ciò che occorre considerare è che le cellule connettivali creano l'ambiente per tutti gli altri tipi di cellule costruendo sia l'impalcatura che le tiene assieme sia la rete di comunicazione fra esse. E’ quindi il cristallo del sistema connettivo a determinare ed evidenziare il nostro stato globale.
Appare chiara l’importanza della ricerca ulteriore in questo campo. Non si può prescindere dallo studio del sistema connettivale se si vuole comprendere a fondo il comportamento globale e locale fisiologico. Lo studio della biochimica non può più essere semplificato in sequenze lineari di reazioni chimico-fisiche ma occorre considerare l’habitat attivo e dinamico in cui la “chimica della vita” si svolge: il sistema connettivo. La stessa psiconeuroendocrinommunologia va quindi estesa alla psiconeuroendocrinoconnettivoimmunologia.
Va infine sottolineato come moderne tecniche e terapie manuali, del movimento e di rieducazione posturale, basate sul concetto di riarmonizzazione del sistema connettivale in accordo con gli altri sistemi, possano avere importanti effetti oltre che immediati anche a lunga durata sulla salute generale dell’organismo.
Fibre della MEC (matrice extracellulare):
 - Fibre Collagene
- Fibre Collagene
Sono le fibre più numerose, impartiscono al tessuto in cui sono presenti colore bianco (ad es. tendini, aponeurosi, capsule degli organi, meningi, cornee ecc.). Formano l'impalcatura di molti organi e sono i componenti più resistenti del loro stroma (tessuto di sostegno). Presentano molecole lunghe e parallele. che si strutturano in microfibrille e quindi in fasci lunghi e tortuosi tenuti assieme da una sostanza cementate contenente carboidrati. Queste fibre sono molto resistenti alla trazione subendo un allungamento del tutto trascurabile.
Le fibre di collagene sono composte principalmente da una scleroproteina, il collagene, proteina di gran lunga più diffusa nel corpo umano rappresentando il 30% delle proteine totali. Questa proteina basica è in grado di modificarsi, in base alle richieste ambientali e funzionali, assumendo gradi variabili di rigidità, elasticità e resistenza. Della sua gamma di variabilità ne sono esempi il tegumento, la membrana basale, cartilagine e l'osso.
 - Fibre Elastiche
- Fibre Elastiche
Queste fibre gialle predominano nel tessuto elastico e quindi in zone del corpo dove è necessaria una particolare elasticità (es. padiglione orecchio, cute). La presenza di fibre elastiche nei vasi sanguinei contribuisce all'efficienza della circolazione del sangue ed è un fattore che ha contribuito allo sviluppo dei vertebrati.
Le fibre elastiche sono più sottili delle fibre collagene, si ramificano e anastomizzano formando un reticolato irregolare, cedono facilmente a forze di trazione riprendendo la loro forma quando la trazione cessa. Il componente principale di queste fibre è la scleroproteina elastina, alquanto più giovane, in termini evolutivi, del collagene.
- Fibre Reticolari
Sono fibre molto sottili (di diametro simile a quello delle fibrille di collagene), considerabili come fibre collagene immature nelle quali in gran parte si trasformano. Sono presenti in grandi quantità nel tessuto connettivo embrionale e in tutte le parti dell'organismo in cui si formano fibre di collagene. Dopo la nascita esse sono particolarmente abbondanti nell'impalcatura degli organi ematopoietici (es. milza, linfonodi, midollo rosso delle ossa) e costituiscono una rete intorno alle cellule degli organi epiteliali (es. fegato, rene, ghiandole endocrine).
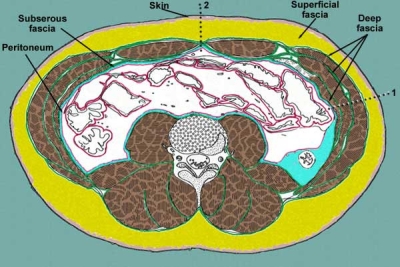 Fra i vari tipi di tessuto connettivo (tessuto connettivo propriamente detto, tessuto elastico, tessuto reticolare, tessuto mucoso, tessuto endoteliale, tessuto adiposo, tessuto cartilagineo, tessuto osseo, a href="circolazione.html#sangue" title="approfondimento" target="_blank">sangue e linfa, la fascia connettivale riveste un particolare interesse dal punto di vista posturale.
Fra i vari tipi di tessuto connettivo (tessuto connettivo propriamente detto, tessuto elastico, tessuto reticolare, tessuto mucoso, tessuto endoteliale, tessuto adiposo, tessuto cartilagineo, tessuto osseo, a href="circolazione.html#sangue" title="approfondimento" target="_blank">sangue e linfa, la fascia connettivale riveste un particolare interesse dal punto di vista posturale.
Prendendo spunto dalla schematizzazione proposta da F. Willard, si può considerare la fascia suddivisa all’incirca in quattro strati formanti cilindri longitudinali concentrici fra loro interconnessi.
1) Lo strato/cilindro più esterno ricoprente tutto il corpo e presente sotto il derma, rappresenta la fascia superficiale. La fascia superficiale è composta da tessuto connettivo lasso (sottocutaneo al cui interno può esserci una trama di fibre collagene e soprattutto elastiche) e adiposo (pertanto il suo spessore, oltre che dalla localizzazione, dipende dalla nostra alimentazione). Tramite fibre, tale fascia forma un continuum con derma ed epidermide verso l’esterno e, al contempo, si ancora ai tessuti e organi sottostanti. La fascia superficiale rappresenta un'importante sede di stoccaggio di acqua e grasso, protegge da deformazioni e insulti meccanici e termici (strato isolante), è una via di passaggio per nervi e vasi sanguigni e permette lo scorrimento della pelle sopra la fascia profonda. Come la fascia profonda presenta poca vascolarizzazione.
2) Sotto la fascia superficiale vi è la fascia profonda, detta anche cervico-toraco-lombare o toraco-lombare, che rappresenta uno strato cilindrico piuttosto coeso intorno al corpo (tronco e arti). Essa è costituita da tessuto connettivo denso irregolare formato da fibre collagene ondulate e da fibre elastiche (disposte secondo andamento trasversale, longitudinale e obliquo) e forma una membrana che riveste la parte esterna muscolare. Questa guaina ricopre il corpo estendendosi dal cranio, a livello del margine della mascella e della base craniale con cui è fusa, da qui si dirige verso gli arti superiori (fino a fondersi con la fascia superficiale a livello dei retinacoli del palmo della mano) e anteriormente passa sotto i muscoli pettorali, ricopre i muscoli intercostali e le coste, l'aponeurosi addominale e si connette alla pelvi. La fascia profonda gira posteriormente connettendosi ai processi trasversi e poi alle apofisi spinose vertebrali formando quindi due comparti (destro e sinistro) contenenti i muscoli paravertebrali.
A livello dell’osso sacro, tale fascia forma un “nodo” inasportabile (in quanto fuso con l'osso) in cui convergono i vari compartimenti fasciali del corpo e da cui si diparte la porzione di fascia profonda che percorre gli arti inferiori fino a fondersi con la fascia superficiale, a livello della pianta del piede nei retinacoli del talo.
Caratteristica distintiva della fascia profonda è quella di formare dei comparti strutturali e funzionali ossia contenenti determinati gruppi muscolari con innervazione specifica. Il compartimento conferisce anche delle caratteristiche morfo-funzionali specifiche al muscolo: un muscolo che si contrae all'interno di una guaina sviluppa una pressione che sostiene la contrazione stessa. I muscoli transversus abdominis costituiscono la parte attiva della fascia toraco-lombare.
A livello del singolo muscolo, la fascia profonda si continua, tramite i setti, le aponeurosi e i tendini, con la fascia muscolare costituita dall'epimisio (tessuto connettivo fibro-elastico che riveste l'intero muscolo) che si estende nel ventre muscolare costituendo il perimisio (tessuto connettivo lasso che riveste i fascicoli di fibre muscolari) e l’endomisio (delicato rivestimento connettivale della fibra muscolare). In condizioni fisiologiche, tali setti e rivestimenti consentono lo scorrimento delle fibre muscolari nonché il loro nutrimento. Questa fascia è direttamente collegata sia anatomicamente che funzionalmente ai fusi neuromuscolari e agli organi tendinei del Golgi.
Come la fascia superficiale, la fascia profonda è scarsamente vascolarizzata e fornisce vie di passaggio per nervi e vasi sanguigni. La fascia profonda (cervico-toraco-lombare) riveste un’enorme importanza posturale (e in particolare nello svolgimento della deambulazione) e di protezione della colonna vertebrale.
Il cilindro costituito dalla fascia profonda contiene due ulteriori cilindri longitudinali posti uno dietro l’altro e formanti, quello anteriore, la fascia viscerale e quello posteriore la fascia meningea.
3) Il cilindro posto anteriormente all’interno della fascia profonda, denominato fascia viscerale o splancnica, è una colonna fasciale che forma il mediastino, estendendosi dalla bocca all'ano tramite varie porzioni con simile struttura ed embriologia: parte dalla base del cranio, si estende giù lungo l'asse mediano (fascia endocervicale, faringea), forma il film ricoprente la pleura parietale dei polmoni (fascia endotoracica), attraversa il diaframma, circonda varie zone della cavità addominale avvolgendo la sacca peritoneale (fascia endoaddominale) e si estende fino alla pelvi (fascia endopelvica). La porzione maggiore di questa fascia si trova intorno agli organi toracici, sull'asse mediano, dove forma una colonna, il comparto mediastinico del torace. Il mediastino toracico si continua quindi con quello addominale fungendo anche da grosso condotto per i fluidi. A livello addominale la fascia endoaddominale si diparte dalla colonna assiale per rivestire completamente gli organi sospesi tornando poi a ricongiungersi con essa (i mesenteri sono ricchi di questa fascia). In alcuni punti la fascia viscerale tende a specializzarsi (ad es. si ispessisce intorno ai reni per proteggerli costituendo la fascia renale).
Questa fascia presenta quindi il grande vantaggio di poter creare degli scomparti ma, essendo anche un deposito di grasso, può creare problematiche di massa deformando la cavità corporea. Ad es. negli obesi può avvenire un'alterazione strutturale e quindi funzionale del diaframma: se l'aumento di massa endotoracico è tale da spingere verso l'esterno le coste, ciò causa un appiattimento del diaframma così che contraendosi, invece di funzionare come muscolo verticale che si abbassa sollevando le coste, traziona i bordi costali verso l'interno trasformandosi in un muscolo espiratorio. In tale situazione sarà impossibile effettuare una fisiologica respirazione profonda e si dovrà ricorrere a respiri brevi, superficiali e frequenti con tutte le conseguenze sulla salute derivanti da ciò.
Alcuni ricercatori considerano questa fascia un tutt’uno con la fascia profonda (cervico-toraco-lombare).
4) Il cilindro posteriore, contenuto nella fascia profonda (cervico-toraco-lombare) e posto dietro la fascia viscerale, rappresenta la fascia meningea che racchiude l'intero sistema nervoso centrale. L'osso craniale, praticamente sospeso sul materiale meningeo, presenta un’origine neuroectodermica sviluppandosi dalla base craniale per differenziazione delle cellule della cresta neurale cranica; esso fa quindi parte dello strato meningeo (e non di quello cervico-toraco-lombare che si ferma, come abbiamo visto, alla base craniale). Asportando l'osso occipitale si accede alla dura madre, punto di partenza superiore della fascia meningea che si estende in giù fino a ca. la II vertebra sacrale tramite il sacco durale. La fascia meningea possiede funzione protettiva e nutritiva del sistema nervoso centrale.
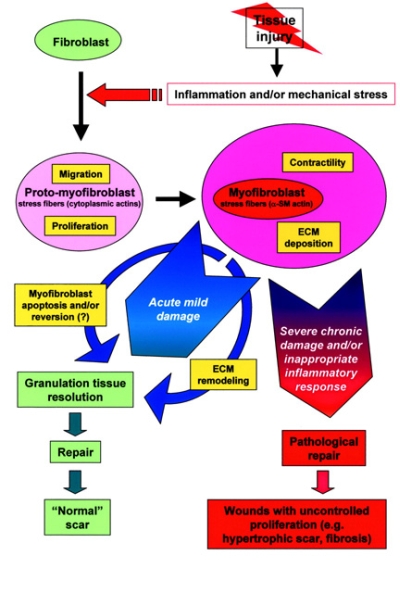 Scoperti nel 1970, i miofibroblasti sono cellule del tessuto connettivo interposte alle fibre collagene fasciali con capacità contrattili simili alla muscolatura liscia (contengono actina). Esse ricoprono un riconosciuto e importante ruolo nella guarigione delle ferite, nella fibrosi dei tessuti e nelle contratture patologiche. I miofibroblasti si contraggono attivamente in situazioni infiammatorie quali morbo di Dupuytren, artrite reumatoide, cirrosi epatica. In condizioni fisiologiche si trovano nella pelle, milza, utero, ovaie, vasi circolatori, setti polmonari, legamenti periodontali. La loro evoluzione è vista generalmente da normali fibroblasti a proto-miofibroblasti, fino alla completa differenziazione in miofibroblasti e a un'apoptosi terminale che è influenzata dalle tensioni meccaniche, dalle citochine e da specifiche proteine che provengono dalla matrice extracellulare.
Scoperti nel 1970, i miofibroblasti sono cellule del tessuto connettivo interposte alle fibre collagene fasciali con capacità contrattili simili alla muscolatura liscia (contengono actina). Esse ricoprono un riconosciuto e importante ruolo nella guarigione delle ferite, nella fibrosi dei tessuti e nelle contratture patologiche. I miofibroblasti si contraggono attivamente in situazioni infiammatorie quali morbo di Dupuytren, artrite reumatoide, cirrosi epatica. In condizioni fisiologiche si trovano nella pelle, milza, utero, ovaie, vasi circolatori, setti polmonari, legamenti periodontali. La loro evoluzione è vista generalmente da normali fibroblasti a proto-miofibroblasti, fino alla completa differenziazione in miofibroblasti e a un'apoptosi terminale che è influenzata dalle tensioni meccaniche, dalle citochine e da specifiche proteine che provengono dalla matrice extracellulare.
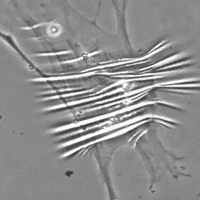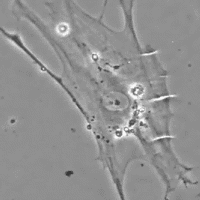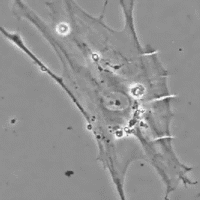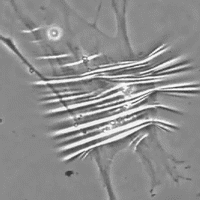 Data anche la favorevole configurazione della distribuzione di tali cellule contrattili all'interno della fascia, il probabile ruolo del miofibroblasta è quello di sistema di tensione accessorio tale da sinergizzare la contrazione muscolare fornendo un vantaggio in situazioni di pericolo per la sopravvivenza (lotta e/o fuga). E' inoltre molto probabile che tramite tali fibre muscolari lisce il sistema nervoso autonomo, tramite nervi intrafasciali, possa “pre-tensionare” la fascia indipendente dal tono muscolare. La presenza di tali cellule nelle capsule di rivestimento degli organi spiegherebbe ad es. come la milza possa rimpicciolirsi fino a metà del suo volume in pochi minuti (fenomeno osservato nei cani in situazioni di sforzo strenuo in cui viene richiesta l’erogazione della scorta di sangue in essa contenuto nonostante che il rivestimento capsulare sia ricco in fibre collagene che consentono solo piccole variazioni di lunghezza.
Data anche la favorevole configurazione della distribuzione di tali cellule contrattili all'interno della fascia, il probabile ruolo del miofibroblasta è quello di sistema di tensione accessorio tale da sinergizzare la contrazione muscolare fornendo un vantaggio in situazioni di pericolo per la sopravvivenza (lotta e/o fuga). E' inoltre molto probabile che tramite tali fibre muscolari lisce il sistema nervoso autonomo, tramite nervi intrafasciali, possa “pre-tensionare” la fascia indipendente dal tono muscolare. La presenza di tali cellule nelle capsule di rivestimento degli organi spiegherebbe ad es. come la milza possa rimpicciolirsi fino a metà del suo volume in pochi minuti (fenomeno osservato nei cani in situazioni di sforzo strenuo in cui viene richiesta l’erogazione della scorta di sangue in essa contenuto nonostante che il rivestimento capsulare sia ricco in fibre collagene che consentono solo piccole variazioni di lunghezza.
La MEC contiene un elevato (e ancora non ben definito) numero di proteine specializzate non collagene che tipicamente presentano domini multipli provvisti di specifici siti di legame per altre molecole della MEC e per i recettori presenti sulla superficie cellulare (fibronectina, laminina, fibrillina ecc.). In questo modo ogni singolo componente di tali proteine funge da “amplificatore” di contatti, sia tra molecole analoghe che diverse, realizzando un infinito network biochimico in grado di generare, modulare, variare e propagare anche a distanza milioni e milioni di informazioni biochimiche (ed energetiche).
Un'importante proteina specializzata della matrice extracellulare è la fibronectina, glicoproteina ad alto peso molecolare rinvenibile in tutti i vertebrati. Essa è un dimero glicoproteico composto da due larghe subunità unite da ponti disulfidrici. La fibronectina appare in grado di influenzare in diversi modi la crescita cellulare, l'adesione intercellulare e con la MEC (possiede la capacità di legarsi alle cellule e ad altre molecole della MEC, es. collagene, fibrina, eparina), la migrazione cellulare (la cellula può spostarsi fino a 5 cm al giorno) ecc. L'isoforma più conosciuto, tipo III, si lega alle integrine. Quest'ultime sono una famiglia di proteine transmembrana che effettuano l'adesione delle cellule alla matrice extracellulare attraverso il riconoscimento della sequenza RGD. I peptidi RGD sono catene aminoacidiche di varia lunghezza a partire da semplici tripeptidi, caratterizzate dalla sequenza aminoacidica Arginina-Glicina-Acido Aspartico. Questa sequenza aminoacidica è presente ubiquitariamente nell’organismo ed è coinvolta in numerose funzioni fisiologiche. Tale sequenza RGD rappresenta una delle strutture che permettono l’adesione di particolari linee cellulari (es. le cellule infiammatorie) così che possano svolgere la loro funzione. Il legame tra integrine e RGD induce una serie di reazioni nel citoplasma che coinvolgono il citoscheletro e altre proteine che regolano l’adesione, la crescita e la migrazione cellulare.
Glucosaminoglicani (GAGs) o glicosaminoglicani (glicosamminoglicani, glicosamminglicani), mucopolisacaridi e proteoglicani (PGs) formano all'interno dei tessuti connettivi una sostanza definita “fondamentale” simil-gel altamente idratata, all'interno della quale le proteine fibrillari risultano ospitate e imbricate. Tale forma di gel polissaccaridico è in grado da un lato di permettere alla MEC di resistere a notevoli forze compressive e dall'altro di consentire una rapida e costante diffusione delle sostanze nutritizie, dei metaboliti e degli ormoni tra sangue e tessuti.
I GAGs si legano di norma covalentemente a un nucleo (core) proteico dando origine ai proteoglicani (PGS). GAGs e PGs sono in grado di agire da soli o in gruppo come recettori per molecole di adesione o come catalizzatori di processi biochimici su molecole circolanti come i fattori di crescita, le citochine e gli enzimi coagulativi.
I glucosaminoglicani (GAGs) sono rappresentati da catene polissacaridiche composte da unità disaccaridiche ripetute numerose volte. Uno dei due zuccheri è sempre rappresentato da un aminozucchero (n-acetilglucosamina o n-acetilgalattosamina) quasi sempre solfato. Il secondo zucchero è di solito l'acido glucuronico o il suo isomero L iduronico. Si riconoscono 4 maggiori gruppi di GAGs: ialuronani, condroitinsolfati e dermatansolfati, eparansolfati, keratansolfati.
Le catene polisaccaridiche dei glucosaminoglicani sono volumetricamente troppo rigide per ripiegarsi all'interno delle compatte strutture globulari tipiche delle catene polipeptidiche, in più esse sono altamente idrofile. Per queste ragioni (e probabilmente anche per altre a noi ignote) i GAGs tendono ad assumere conformazioni estremamente estese occupando un grande volume in rapporto alla loro massa e formando così quantità notevoli di gel anche a basse concentrazioni. L'elevata quantità di cariche negative (i GAGs rappresentano le più numerose cellule anioniche, essendo di norma solfate, prodotte dalle cellule animali) attrae numerosi cationi; tra questi un ruolo predominante lo svolge il Na+ che conferisce all'insieme capacità osmotiche e intrappola un'enorme quantità di acqua nella MEC. In tal modo si generano rigonfiamenti (turgori) che consentono alla MEC di opporsi a forze compressive anche importanti (grazie a ciò, ad es., la cartilagine dell'anca può, in condizioni fisiologiche, resistere perfettamente a una pressione di diverse centinaia di atmosfere).
All'interno del tessuto connettivo, i GAGs rappresentano meno del 10-12% del peso globale, essi però, grazie alle loro caratteristiche, riempiono molti degli spazi extracellulari formando dei pori di gel idratato di varie dimensioni e densità di cariche elettriche fungendo così da punti chiave selettivi o “server” attraverso i quali viene regolato il traffico di molecole e cellule all'interno della MEC, in base alle loro dimensioni, peso e carica elettrica.
L'acido ialuronico (ialuronano, ialuronato) rappresenta forse il più semplice dei GAGs. Nell'uomo è costituito da circa 25.000 disaccaridi uguali non solfati e non si mostra di solito collegato ad alcun “core proteico”, pertanto presenta una struttura atipica. Dati sperimentali e biologico-molecolari confermano che esso gioca un ruolo fondamentale a livello delle ossa e delle articolazioni riguardo la resistenza a notevoli pressioni. Inoltre ricopre un importantissimo compito di riempimento di spazi nella MEC durante lo sviluppo embrionale: crea spazi vuoti fra le cellule in cui le stesse, in fasi successive, migreranno.
GAGs e PGs associandosi tra loro possono dare origine ad enormi complessi polimerici nella MEC. Ad esempio le molecole di aggrecano, che rappresentano la maggioranza dei PGs a livello articolare, si combinano tramite legami non covalenti con l'acido ialuronico dando origine ad aggregati delle dimensioni di un batterio.
Non tutti i PGs sono secreti dalla MEC, alcuni sono componenti integrali delle membrane plasmatiche. Tra i PGs di membrana più conosciuti, i sindecani presentano un dominio extracellulare costituito da tre catene di GAGs mentre quello intracellulare è ritenuto in grado di reagire con l'actina del citoscheletro cellulare.
- Il Sistema Connettivo, Giovanni Chetta
- Postura e Benessere, Giovanni Chetta
- La Scoliosi quale atteggiamento naturale, Giovanni Chetta
- Dalla MEC alla postura, Giovanni Chetta
- Massaggio & Bodywork TIB, Giovanni Chetta
- Ginnastica Posturale TIB, Giovanni Chetta
- Alimentazione e Benessere, Giovanni Chetta
- Stress e Benessere, Giovanni Chetta
